
第四章 免疫系统
免疫系统(immunesystem)是由具有免疫功能的器官、组织、细胞和分子组成,是机体免疫机制发生的物质基础。免疫系统内的各种淋巴样器官和细胞在机体的整体免疫功能中分别担负着不同的角色,根据其功能不同可将整个系统分成3个组织层次:①中枢免疫器官;②外周免疫器官;③免疫细胞。各层次不同类型的组织与细胞又有着不同的作用,通过淋巴细胞再循环和各种免疫分子将各部分的功能协调统一起来。与机体的其它系统一样,免疫系统虽有着一系列的内部调节机制,但不是完全独立运行,而是与其它系统互相协调,尤其是受神经体液调节,又可进行反馈影响,共同维持机体的生理平衡。
免疫系统是伴随着生物种系发生和发展过程中逐步进化而建立起来的。无脊椎动物仅有吞噬作用和炎症反应,到了脊椎动物才开始有腔上囊,出现特异性抗体,至哺乳动物才逐渐产生较多种类的免疫球蛋白。进化程序不同的动物中免疫球蛋白类型出现的多少不一。免疫系统各成分的系统发生顺序为吞噬细胞、细胞介导免疫、体液免疫;在体液免疫中抗体产生的顺序是IgM、IgG、IgA、IgD和IgE(图4-1)。
图4-1免疫应答的种系进化示意图
第一节 免疫器官
免疫器官(immuneorgan)是指实现免疫功能的器官或组织。根据发生的时间顺序和功能差异,可分为中枢神经免疫器官(centralimmuneorgan)和外周免疫器官(peripheralimmuneorgan)两部分。
一、中枢免疫器官
中枢免疫器官又称一级免疫器官,包括骨髓、胸腺、鸟类法氏囊或其同功器官。中枢器官主导免疫活性细胞的产生、增殖和分化成熟,对外周淋巴器官发育和全身免疫功能起调节作用。
(一)胸腺
1.胸腺的组织结构胸腺位于前纵隔、胸骨后。胸腺分为左右两叶,外包结缔组织被膜;被膜伸入胸腺实质内形成隔膜,将胸腺分成许多小叶;小叶的外周部分称为皮质,中央部分称为髓质;相邻的小叶髓质彼此相连(图4-2)。
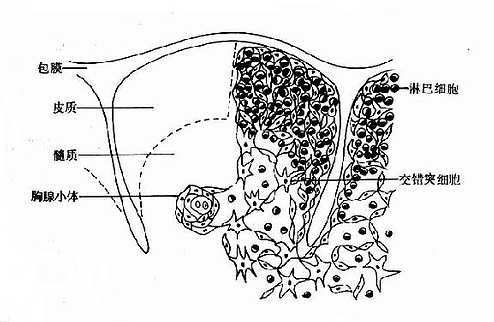
图4-2胸腺结构示意图
胸腺的细胞分为淋巴样细胞和非淋巴细胞两类。淋巴细胞包括原始T细胞向成熟T细胞分化过程中各种不同阶段的细胞,统称为胸腺细胞;胸腺细胞是胸腺内的主体细胞,其分布从皮质到髓质逐渐减少。非淋巴细胞包括上皮细胞、巨噬细胞、树突状细胞、抚育细胞、皮纤维细胞和网状细胞等。这些细胞一方面构成胸腺组织的支架,另一方面构成胸腺细胞营养和分化的微环境,统称为基质细胞。
胸腺皮质的毛细血管内皮细胞连接紧密,与网状细胞共同形成血液-胸腺屏障,使循环中的抗原物质不能进入胸腺。血液-胸腺屏障是体内为数不多的几个生理屏障之一,其意义目前尚不清楚。胸腺髓质的毛细血管内皮细胞之间有间隙,抗原性物质可进入髓质,在髓质内还可见多层扁平上皮细胞呈同心圆状排列成的Hassall小体,或称胸腺小体。直径约25~50μm,其功能尚不清楚。
2.胸腺的免疫功能长期以来对胸腺的功能不甚了解,直到60年代初Miller和Good分别用切除新生小鼠和家兔胸腺的办法证明了胸腺的免疫功能。
(1)训化T细胞:在骨髓初步发育的淋巴细胞经由血液循环迁移至胸腺,定位于胸腺的皮质外层;这些形体较大的细胞为双阴性(CD4-/CD8-)细胞,约占胸腺细胞总数的10%。外层细胞在胸腺微环境中迅速增殖,并推动细胞不断向内层迁移,个体形态逐渐变小;内层细胞为双阳性(CD4+/CD8+)细胞,约占胸腺细胞总数的75%。双阳性细胞为过渡态细胞,其中90%以上在皮质内凋亡或被巨噬细胞吞噬;据认为,死亡细胞可能是针对自身抗原进行应答的细胞。少数胸腺细胞继续发育并迁移至髓质,成为单阳性(CD4+或CD8+)细胞,约占胸腺细胞总数的15%。只有这些单阳性细胞才是成熟的T细胞,通过髓质小静脉进入血循环。
(2)分泌胸腺激素:胸腺上皮细胞能产生多种激素,如胸腺素、胸腺生成素和胸腺体液因子等。这些激素可以诱导活化未成熟胸腺细胞的末端脱氧核苷转移酶,促进T细胞的分化成熟;不同的激素作用于不同的细胞发育阶段,有选择地发挥免疫调节功能。胸腺激素的作用没有种属特异性,所以目前临床应用的胸腺素都是从动物胸腺中提取出来的。
(3)其他:胸腺还可促进肥大细胞发育,调节机体的免疫平衡,维持自身的免疫稳定性。新生动物摘除胸腺,可引起严重的细胞免疫缺陷和总体免疫功能降低。由此可见胸腺在免疫系统中的地位。
3.胸腺的发育过程胸腺于胚胎第6周时就在第三对咽囊的腹侧面形成胚基,至第7周形成胸腺雏形,至第20周时便已发育成熟。出生时胸腺重量仅约为20g,青春期达顶峰,约40g;以后随年龄增长而逐渐萎缩,至老年时仅剩10g左右,且多为脂肪组织替代。机体的免疫功能与胸腺的生长周期相关。
(二)腔上囊
腔上囊又称法氏囊(bursaofFabricius),是鸟类动物特有的淋巴器官,位于胃肠道末端泄殖腔的后上方。与胸腺不同,腔上囊训化B细胞成熟,主导机体的体液免疫功能。将孵出的雏鸡去掉腔上囊,会使血中γ球蛋白缺乏,且没有浆细胞,注射疫苗亦不能产生抗体。
人类和哺乳动物没有腔上囊,其功能由相似的组织器官代替,称为腔上囊同功器官;曾一度认为同功器官是阑尾、扁桃体和肠集结淋巴结,现在已证明是骨髓。
(三)骨髓
骨髓是成年人和动物所有血细胞的唯一来源,各种免疫细胞也是从骨髓的多能干细胞发育而来。
骨髓的主要功能是产生血细胞,近来证明骨髓还是腔上囊同功器官。在骨髓异常时,累及的不单是体液免疫,其他免疫功能也发生障碍。
二、外周免疫器官
外周免疫器官包括淋巴结、脾和粘膜相关淋巴组织(mucosaassociatedlymphoidtissue,MALT)等,是免疫细胞聚集和免疫应答发生的场所。
(一)淋巴结
1.淋巴结的结构淋巴结为近乎圆形的网状结构,表面有一层结缔组织被膜,略凹陷处为门,有输出淋巴管和血管出入。被膜向外延伸有许多输入淋巴管;向内伸入实质形成许多小梁,将淋巴结分成许多小叶。淋巴结的外周部分为皮质,中央部分为髓质(图4-3)。
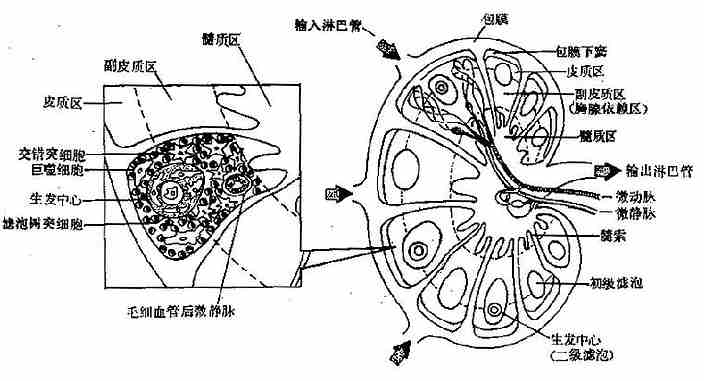
图4-3淋巴结结构示意图
皮质区有淋巴小结,又称淋巴滤泡;受抗原刺激后出现生发中心;此区内富含B细胞和滤泡树突状细胞(follicledendritic,FDCs),所以又称非胸腺依赖区。皮质深层和滤泡间隙为副皮质区,因富含T细胞又称胸腺依赖区;此区是淋巴细胞再循环的门户,有大量T细胞和巨噬细胞分布在滤泡周围,是传递免疫信息的场所。髓质区的B细胞、浆细胞和网状细胞集结成索状,称髓索;在髓索这间为髓窦;此区是滤过淋巴液的场所。
2.淋巴结的功能
(1)滤过和净化作用:淋巴结是淋巴液的有效滤器,通过淋巴窦内吞噬细胞的吞噬作用以及体液抗体等免疫分子的作用,可以杀伤病原微生物,清除异物,从而起到净化淋巴液,防止病原体扩散的作用。
(2)免疫应答场所:淋巴结中富含各种类型的免疫细胞,利于捕捉抗原、传递抗原信息和细胞活化增殖。FDCs表面有丰富的Fc受体,具有很强的捕获抗原体复合物的能力,通过这种方式可将抗原长期保留在滤泡内,这对形成和维持B记忆细胞、诱导再次免疫应答很有意义。B细胞受刺激活化后,高速分化增殖,生成大量的浆细胞形成生发中心;T细胞也可在淋巴结内分化增殖为致敏淋巴细胞。不管发生哪类免疫应答,都会引起局部淋巴结肿大。
(3)淋巴细胞再循环基地:正常情况下,只有少数淋巴细胞在淋巴结内分裂增殖,大部分细胞是再循环的淋巴细胞。血中的淋巴细胞通过毛细血管后静脉进入淋巴结副皮质,然后再经淋巴窦汇入输出淋巴管。众多的淋巴结是再循环细胞的重要补充来源。
(二)脾
1.脾的组织结构脾是体内形体最大的淋巴器官,结构类似淋巴结。脾的表面有结缔组织被膜,实质比较柔脆,分为白髓和红髓。白髓是淋巴细胞聚集之处,沿中央小动脉呈鞘状分布,富含T细胞,相当于淋巴结的副质区。白髓中还有淋巴小结,是B细胞居留之处,受抗原刺激后可出现生发中心。脾中T细胞约占总淋巴细胞数35%~50%,B细胞约占50%~65%。红髓位于白髓周围,可分为脾索和血窦。脾索为网状结缔组织形成的条索状分支结构;血窦为迂曲的血管,其分支吻合成网。红髓与白髓之间的区域称为边缘区,中央小动脉分支由此进入,是再循环淋巴细胞入脾之处。与淋巴结不同,脾没有输入淋巴管,只有一条平时关闭的输出淋巴管与中央动脉并行,发生免疫应答时淋巴细胞由此进入再循环池。
2.脾的功能脾在胚胎期是重要的造血器官;出生后造血功能停止,但仍然是血细胞尤其是淋巴细胞再循环池的最大储库和强有力的过滤器;①与淋巴结相似,脾还是发生免疫应答的重要基地。此外,脾还有两个显著的特点:产生抗体,脾富含B细胞和浆细胞,因此是全身最大的抗体产生器官,尤其是产生IgM和IgG,其数量对调节血清抗体水平起很大作用。所以当自身抗体产生过多导致严重疾病时,曾用切除脾的办法进行缓冲治疗;但脾切除后机体的抗感染能力显著降低。②分泌体液因子,脾可以合成补体(C5和C8等)和备解素等重要的免疫效应分子;还能产生一种白细胞激肽,促进粒细胞的吞噬作用。
(三)粘膜相关淋巴组织
在各种腔道粘膜下有大量的淋巴组织聚集,称为粘膜相关淋巴组织(MALT);其中最重要的是胃肠道粘膜相关淋巴组织(GALT)和呼吸道粘膜相关淋巴组织(BALT)。GALT包括阑尾、肠集合淋巴结和大量的弥散淋巴组织;BALT包括咽部的扁桃体和弥散的淋巴组织,构成呼吸道和消化道入口处的防御机构,称为Waldeyer环。除了消化道和呼吸道外,乳腺、泪腺、唾液腺以及泌尿生殖道等粘膜也存在弥散的MALT。
与淋巴结和脾不同,粘膜相关淋巴组织没有包膜,不构成独立的器官,通过广泛的直接表面接触和体液因子与外界联系;MALT中的B细胞多为IgA产生细胞,受抗原刺激后直接将SigA分泌到附近粘膜,发挥局部免疫作用;粘膜靠一种特殊的机制吸引循环中的淋巴细胞,MALT中的淋巴细胞也可输入到淋巴细胞再循环池,某一局部的免疫应答效果可以普及到全身的粘膜(详见第七章)。
三、淋巴细胞再循环
各种免疫器官中的淋巴细胞并不是定居不动的群体,而是通过血液和淋巴液的循环进行有规律的迁移,这种规律性的迁移为淋巴细胞再循环(lymphocyterecirculation)。通过再循环,可以增加淋巴细胞与抗原接触的机会,更有效地激发免疫应答;并不断更新和补充循环池的淋巴细胞。
1.再循环的细胞淋巴干细胞从骨髓迁移至胸腺和腔上囊或其功能器官,分化成熟后进入血液循环的定向移动过程不属于再循环范围。再循环是成熟淋巴细胞通过循环途径实现淋细胞不断重新分布的过程。再循环中的细胞多是静止期细胞和记忆细胞,其中80%以上是T细胞。这些细胞最初来源于胸腺和骨髓;成年以后,再循环池手细胞主要靠外周免疫器官进行补充。受抗原刺激而活化的淋巴细胞很快定居于外周免疫器官,不再参加再循环。
2.再循环的途径血液中的淋巴细胞在流经外周免疫器官(以淋巴结为例)时,在副皮质区与皮质区的连接处穿过高内皮毛细血管后静脉(HEV)进入淋巴结;T细胞定位于副皮质,B细胞主要定位于皮质区;以后均通过淋巴结髓窦迁移至输出淋巴管,进入高一级淋巴结;经过类似的路径,所有外周免疫器官输出的细胞最后都汇集于淋巴导管;身体下部和左上部的汇集到胸导管,从左锁骨下静脉角返回血循环;右侧上部的汇集到右淋巴管,从右锁骨下静脉返回血循环(图4-4)。再循环一周约需24~48小时。
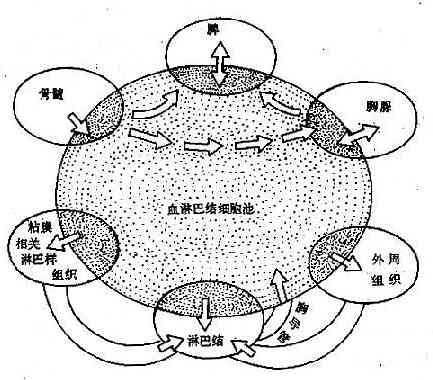
图4-4体内淋巴细胞迁移路线示意图
3.细胞定居选择淋巴细胞从血循环进入淋巴组织具有高度的选择性,这是因为淋巴细胞上具有特殊的受体分子,称为归巢受体(homingreceptor)。现已发现的归巢受体包括CD44、LFA-1、VLA-4和Mel-14/LAM-1等;其中Mel-14/LAM-1是定居淋巴结的受体,识别淋巴结内的高内皮细胞;VLA-4的α亚单位是定居MALT的受体,识别粘膜表面的配体。