四、遗传工程与近交系的发展
遗传工程是70年代兴起的一门崭新的科学技术,它使人类进入了定向控制生物遗传性状的新阶段。
所谓遗传工程也就是根据人们的意愿,采用工程建筑的手法,按照预先设计的方案,借助于实验室手段将一个生物体的遗传物质定向地转移到另一个生物体中去,使后者获得人类希望的性状,成为一个新的“物种”。这们就打破了传统的、必须经过两性杂交的育种方法,使原来在自然状态下根本不可能发生有性繁殖的两种生物的基因结合在一起了。
遗传工程的概念可分为广义的和狭义的两种。广义的遗传工程包括细胞工程、染色体工程和基因工程三类,使用的是细胞生物学方法和分子生物学方法,包括细胞融合、细胞拆合、染色体导入和基因或DNA分子的转移等。广义的遗传工程又称为微生物工程或细胞工程。狭义的遗体工程仅限于基因工程。
早期的遗传工程的研究工作大多数是在病毒、噬菌体和细菌中进行的。因为这些生物的遗传物质比较简单,容易搞清楚遗传物质与生物性状之间的关系。随着知识的积累,技术的提高,现在越来越多的遗传工程实验开始在高等哺乳类动物身上进行了。这尤其促进了近交系小鼠的发展。
(一)细胞工程与近交系小鼠的发展
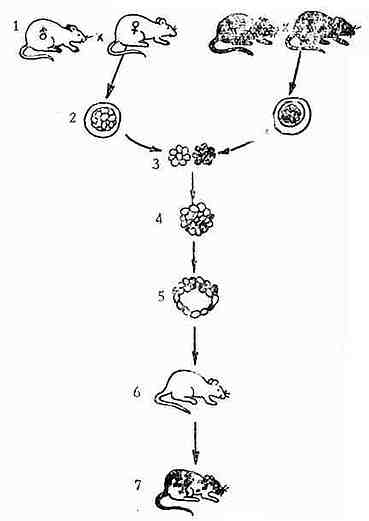
1.嵌合体小鼠(Mouse Aggregation Chimeras)的育成。嵌合体小鼠技术是由Tarkowski和Mintz分别在华沙和费城发展的(见图3-5)。他们首先用两对不同品系的近交系小鼠在同一时刻进行纯交,为了使其后代便于区分,常选择毛色不同品种的近交系小鼠例如选用白毛的SJL小鼠与黑毛色的BL/10小鼠。当受精卵分裂为8分裂球时,分别将它们从各自母体的子宫上分离下来。然后用蛋白酶消化分裂球外面的明带,使分裂球“裸露”。并在37℃的条件下,将来自两个品种的两个分裂球彼此接触,任其粘成一个具有双倍体积的早期胚胎。将早期胚胎继续培养到具有128~256个细胞的胚囊。这时,不同毛色品系的细胞相互混杂发育在一起。然后,通过手术把胚囊移植到寄养母鼠的子宫内,让它继续发育,直至出生。新生小鼠长出毛后,其毛色表现为既不象父亲SJL品系的全白色,也不象母系BL/10品系的全黑色,而是表现出黑、白条或块状的毛色。这说明新生鼠的组织是由“黑色”细胞和“白色”细胞嵌合而成的,是一只嵌合体小鼠。
图3-5 嵌合小鼠育成
近交同类系动物也是近交系动物,它除了一小段带有可辩的目的基因染色体外,在遗传上与原来已建立起来的那个近交系完全一致。近交同类系动物的培育,是选用带有目的基因的个体与已经建立起来的近交系杂交建立起的。应用近交同类系动物可以研究多基因系统中一个基因的特殊作用,施耐尔博士(Snell)是第一个应用这种体系进行组织移植基因研究的,建立了同类抵抗系学说,为小鼠的主要组织相容性抗原的研究做出了巨大贡献,为此获得了1981年的诺贝尔奖金。
同类系动物的命名基于如下两种方式。一种是带有近交品系和供体品系的复合符号,中间带有一点,例:B6・AKR。第二种命名法是近交品系后面加上一横,然后标上供体品系特异的基因位点的符号。这样B6・AKR也可以写作B6-H-2k。对于最近培育的同类系,小鼠遗传标准命名委员会建议同时采用两种方式,这样B6・AR或B6-H-2k应该写为B6・AKR-H-2k。
如果进一步研究嵌合体小鼠的其它性状,还可以发现嵌合体小鼠将双亲的各种不同的性状都嵌合起来了。例如Mintz(1967,1971)证明用DBA/2和C3H两种不同品种的近交系所嵌合的后代具有不同的异柠檬酸脱氢酶(IDH-1)或苹果酸脱氢酶(MDH),利用电泳技术可以将它们彼此分开。除了双亲具有的酶以外,嵌合体内有时还可以发现第三种酶,即双亲的“杂种”或异多聚酶。以上的工作对于细胞水平的遗传性研究是具有重要意义的,受到人们越来越多的重视。
嵌合体小鼠近年来已应用于细胞和组织的动力学研究,如研究小肠上皮细胞移行规律及其定位等。
2.单亲纯合双倍体动物的育成。单亲纯合双倍动物育成技术又称为雌核发育技术。这是一种相当于植物中由花粉培育成纯合双倍体植株的技术,其结果都是培育出具有两套完全一致的染色体及基因的个体。
众所周知,不论在生产上还是在科学试验中都需要纯系动物。为了得到一个纯系动物,一般要花费数年时间,甚至一生的光阴。既使对世代周期较短的小鼠,采用了最有效的兄妹交配方法,还需要连续兄妹交配20代以上才能获得纯系动物,这也要几年时间。如果采用单亲纯合双倍体育成的技术来育成一个纯系动物,则将大大地缩短育种周期,小鼠只需三周,牛也只需几个月。具体方法见图3-6。
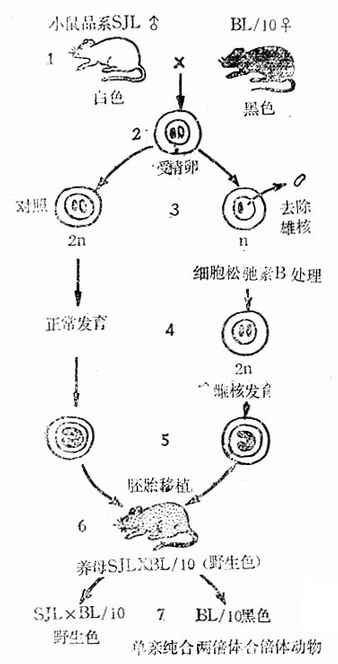
图3-6 单亲双位体动物育成(雌核发育)
首先将两个不同品系的的近交系进行杂交(例如SJLXBL/10),交配后,在精核与卵核尚未融合之前,从母鼠子宫内冲取受精卵并用极细的吸管将雄核去掉。然后在细胞松驰素B的处理下使雌核加倍,形成二倍体细胞,二倍体细胞在体外继续培养到胚囊期后,移植到养母的子宫内使胚胎继续发育,直至出生。一般选择毛色与双亲不同的品系作为养母,以易于区分。本例中是选择毛色为野生色的BL/10XSJL杂交鼠作为养母的。养母本身没有经过交配,因而出生的小鼠均为移植进去的胚胎发育而成,它表现出亲母鼠纯合的性状。若出生的新生鼠表现为野生色,则说明实验技术失败,并未剔除精核。
单亲纯合双倍体技术也已在鱼类和家兔中获得成功。
3.细胞核移植系的产生。在单亲纯合双倍体育成技术中已提到了核移植技术,但这仅是该技术的初步。若能将受精卵中的精核,与卵核都移出,然后再将来自其它细胞新核移入受精卵,并能顺利地完成新胚胎的正常发育,核移植技术就算成功了。以前,完整的核移植技术仅在两栖类和昆虫中取得成功。1981年日内瓦大学的伊尔曼斯(K.Illmensee)和美国杰克逊研究所的霍普(P.C.Hoppe)首次在小鼠中完成了核移植技术,开创了哺乳动物核移植成功的先例。具体方法见图3-7。
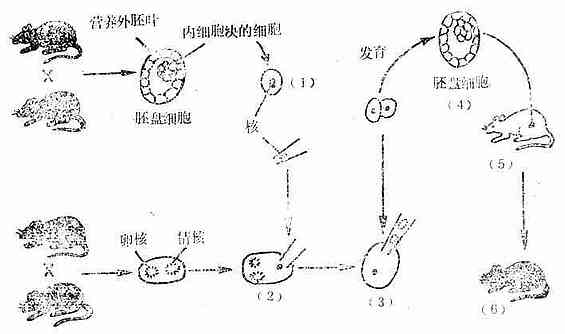
图3-7细胞移植系的产生
(1)将内细胞块细胞里的核用吸管吸出。
(2)将取出的核输到另一个精核与卵核尚未融合的受精卵内。
(3)吸出该受精卵本身的精核与卵核。
(4)受精卵在体外发育到胚胎细胞。
(5)将胚盘细胞移植到白色的养母小鼠的子宫内。
(6)出生的小鼠长出灰毛,说明细胞移植系实验成功。
(1)首先用灰色的小鼠品系进行雌雄交配(也可用其它毛色的品系),从母体的子宫内获得胚盘细胞。胚盘细胞包括营养外胚叶细胞和内细胞块细胞两种,后者将来可形成胚胎。若错误地取了营养外胚叶细胞就要导致实验失败。
(2)用极细的吸管取出内细胞块细胞的细胞核,并将它注到刚受精的、但卵核与精核尚未融合的黑色小鼠的受精卵内。
(3)从黑色小鼠的受精卵内取出吸管时,将黑色鼠受精卵原有的卵核和精核吸出。
(4)将处理过的受精卵在体外培养到胚盘细胞期,再将它移植到白色的养母小鼠的子宫内,让它继续发育。
(5)产出的新生小鼠必须会长出灰色的毛,这就是细胞核移植鼠。
由于主要的遗传物质存在于细胞核中,所在核移植的受体将具有与核移植供体完全相同的遗传基础。因此,细胞核移植系技术在动物育种工作中具有十分重要的意义。因为它象植物中的组织培养那样,可以利用一头优质的动物(例如高产的奶牛)复制出与它生得完全一样的成千上万的细胞核移植系动物。
(二)实验小鼠染色体工程进展
所谓染色体工程是以染色体为单位进行有意识的切割、修补或成条、成套地增加,将人类需要的遗传性状集中在一起,创造出新的物种。以往的染色体工程一般均采用物理的或化学的方法。在高等动物中,由于多倍体或者染色体严重缺失与重复的个体都不容易成活,所以高等哺乳动物的染色体工程目前均是在细胞水平上采用细胞融合技术进行。
Harris和Watkins(1965)首先制备了包含小鼠和人类细胞的种间异核本。他们将由宫颈癌组织培养出来的人类HaLa细胞和小鼠细胞(生长在腹膜腔中Ehrlich小鼠肿瘤细胞)置于同一培养液中,加入经紫外线灭活了的仙台病毒(Sendai Virus,是粘病毒的副流感群中的一种)。在仙台病毒的作用下使两种细胞发生融合。融合细胞开始有两个来自不同物种的细胞核,称为异核体,这一点可以通过同位素标记的方法加以证实,事先他们用氚标记的胸苷来标记HaLa细胞的细胞核,而在一个异核体细胞内,可以发现标记的和不标记的两个细胞核。异核体的两个核融合后就可以继续进行细胞分裂,并形成单核细胞系。杂种细胞大,容易认别。它在以后的不断的分裂过程,人类的一套染色体和小鼠的一套染色体并不是都随着细胞的分裂进行复制的。在正常的条件下,人类的染色体将随着细胞的分裂而逐渐丢失。这种丢失是随机的。所以最后形成的杂种细胞往往都是带有一整套小鼠染色体加上不同数目和不同编号的人类染色体(或带有丝粘的片断),由于现代染色技术发展,使人们不仅可根据染色的结果很容易地区分人类染色体与小鼠染色体,而且可以确定在杂种细胞中留下来的是哪一条或者哪几条人类染色体。
如果使用的小鼠细胞是营养缺陷型细胞,即细胞本身不能合成某种必需营养物,只有在培养液中添加了这种营养物质后细胞才能正常地生长。小鼠营养缺陷型根据其所需的不同种营养物有许多不同的种类。如果将某种小鼠营养缺陷型细胞与人类正常细胞融合,形成杂种细胞。这时小鼠的营养缺陷可由于人类染色体上的有关基因的存在,该种生存必需营养物得到弥补,杂种细胞在不添加该营养液中的也能正常生长。经过一段时期培养,保留下来的细胞除带有一整套小鼠染色体外,至少还稳定地保留了一条与该营养物合成有关的人类染色体。如果我们使用多种营养缺陷型细胞作为遗传标志,就可有得到许多不同的杂种细胞,通过分析比较,就可能确定在人类的遗传中那种营养物质合成的基因在哪一条染色体。我们不仅可以用营养缺陷基因作为标志,还可以采用生化、免疫学和医学有关的基因作为标志基因,这样就可以把决定嘌呤酶的基因,决定各种碳水化合物、氨基酸以及脂肪代谢的基因,氨基酰-tRNA合成的基因,遗传性疾病的基因(例如白内障、指甲髌骨发育不全),对白喉和脊髓灰质炎等毒素易感性基因,细胞表面抗原基因和多肽酶基因等作为标志基因,结果就可以完成对更多的基因染色体定位工作。现在已定位在人类染色体的基因数目已超过了210个,对人类遗传学的研究作出了很大的贡献。
(三)实验小鼠基因工程进展
超级小鼠的产生。1982年末Palmiter和Brister报导,他们把小鼠MT-1基因的启动子与大鼠生长激素(GH)基因结合,制成融合基因(MGH),然后将MGH插入到大肠杆菌的质粒中,扩增,备用;把收获的MGH基因注入小鼠受精卵的雄性前核;然后将这些受精卵移植到养母小鼠的子宫内,直至分娩。
结果在出生的21只小鼠中有7只带有MGH基因,6只小鼠的每个细胞均有两个以上的MGH(多的可达35个)。在携带MGH的小鼠肝细胞中检出的MGH-mRNA数目高达3000个血液中生长激素浓度明显增高,表明MGH基因已得到表达。含有两个以上MGH基因的6只小鼠,生长迅速,个别的为正常小鼠的两倍,成为所谓的“超级小鼠”。含有MGH基因的小鼠与普遍小鼠杂交的结果,子代19只小鼠中有10只含有MGH基因。MGH基因可以从亲代向子代递这一事实说明MGH基因已整合到小鼠染色体上去了。
最近也有人报导了利用人类的生长激素基因使小鼠成为超级小鼠的。超级小鼠的育成,开创了基因工程在哺乳类动物中获得成功的首例。它对于医学和畜牧业的发展有着极其深远的意义和广阔的前景。