�ڶ�ʮ���¡�DNA���鼼�������ڻ�������е�Ӧ��
��һ�ڡ�����ø
�������̵Ļ����������˹����л���ļ��С�ƴ�ӡ���ϡ�������һ�ξ���һ�����ܵ�DNA���ӣ�Ҫ�Ѳ�ͬ�����DNA���η���Ƭ��ȷ���г�������Ҫ���������Ժ�������ø��restriction endonuclease����Ҫ�Ѳ�ͬƬ��������������ҪDNA����ø��ligase����Ҫ��ϻ�������е�һ��Ƭ�Σ���ҪDNAø��DNa polymerase���ȡ���ˣ�ø��DNA���鼼���бز����ٵĹ��ߣ���������Ҫ�õ�øͳ��Ϊ����ø��
����һ��DNA����������ø
����Lurva��Human��1952���Լ�Bertani��Weigle��1953���������ɾ���˵��������ã�����һ�֦��ɾ�����һ������ϸ���������ã�������һ������ϸ���������ܲ��ԭ����������DNA�ܵ���һ�������ġ����ơ����ɴ˷���������-����ϵͳ��
��������ϸ�����ܺϳ�һ�ֻ���˳��רһ�ĺ�������ø����Щø�и�DNA��˫������Ϊ���ǵĹ��ܾ����и�DNA��������Դ��DNA����������ϸ���ڣ����Գ����ֺ�������øΪ����ø���ϳ�����ø��ϸ��������DNA���Բ�������ø�����ã���Ϊϸ�����ϳ���һ������ø�����ı�������øʶ���DNA˳��Ľṹ��ʹ����ø���������á�����-����ϵͳ��ϸ����һ�ַ����ֶΡ�������ɾ���ȥ��Ⱦ����-����ϵͳ�л��Ե�ϸ�����ɾ���DNAû���Ⱦ����Σ������Ⱦ����ε��ɾ�����ȣ���ȾЧ��Ҫ�ͼ�����������δ�����ε��ɾ���DNA����ϸ��������ø�г�Ƭ�Σ�Ƭ�ε���Ŀ��DNA����������ø��ʶ�����Ŀ�����ȣ���ЩƬ�ν�һ����ϸ���ĺ�������ø���⣬�ͻῪʼ�ѽ��Ⱦ���ɴ˲������Ӵ��ɾ���ȫ���������ι���DNA��������Ժܸߵ�Ч��ȥ��Ⱦ��һЩ������ͬ����-����ϵͳ��ϸ����Ŀǰ���Ӹ��������з����������������ø�ѳ���175�֣�����80�������и�DNA˫����
������һ������ԭ��
��������������ø��Ҫ�Ǵ�ԭ����������ȡ�ġ�����ͨ�õ�����ԭ���ǣ���һ������ϸ�������ĵ�һ����ĸ���ڶ�����������ϸ��������ǰ������ĸ����Щ��ĸ����б����ĸ������ȥ��ϸ����ĵ�һ����ĸ����������ĸ��д�����ͬһ�������м��ֲ�ͬ������øʱ����ֱ����������֢��������������б�����˵�����£���23-1����
��23-1 ��������������ø����ԭ�����
| ϸ��ԭ�� | ϸ������ | �������� | ����ø���� |
| Arthrobacter | Luteus | �� | Alu�� |
| Bacillus | amyloliquefaciens | H | BamH�� |
| Escherichia | Coli | RY13 | EcoR�� |
| Haemophilus | influeuzae | Rd | Hind�� |
�������������������
��������������ø��Ҫ�ֳ������ࡣ��һ������������ø��ʶ��רһ�ĺ�����˳����ʶ��㸽����һЩ���������и�DNA�����е�˫���������и�ĺ�����˳��û��רһ�ԣ�������ġ���������������ø��DNA���鼼���������û�ж���ô��������ڷ���DNA�ṹ���¡��������ø��EcoB��EcoK�ȡ�
�����ڶ�������������ø��ʶ��רһ�ĺ�����˳���ڸ�˳���ڵĹ̶�λ�����и�˫����������������������ø��ʶ����и�ĺ����ᶼ��רһ�ġ��������ܵõ�ͬ��������˳���DNAƬ�Σ����ܹ������Բ�ͬ�������DNAƬ�Σ��γ��Ӻ�DNA���ӡ���ˣ���������������ø��DNA���鼼������õĹ���ø֮һ������øʶ���רһ������˳���������4����6�������ᣬ����Ҳ��ʶ��5���������Լ�7����9����10����11��������ġ����ʶ��λ����DNA�����зֲ�������ģ���ʶ��4�������������������øÿ��46��4096�������������һ���е㡣�˵ĵ����������ݹ���Ϊ3��199�����ᣬʶ��4�������������������ø���е㽫�У�3��109/2.5��102��Լ107���е㣬Ҳ���ǿɱ�����ø�г�107Ƭ�Σ�ʶ��6�������������������øҲ���У�3��109/4��103��Լ106���е㡣
�����ڶ�������������ø��ʶ��˳����һ�����ĶԳ�˳����һ�����ĶԳ��ᣬ������ᳯ��������������ȫ��ͬ������ø���и���������ַ�ʽ��һ�ǽ����и����γ���������ĩ�ˣ�����ĩ�˵ĺ�����˳���ǻ����ģ����γ���������Գ�Ϊճ��ĩ�ˡ���EcoRI��ʶ��˳��Ϊ��
�������������������������������������� |
5������GAA | TTC����3��
3������CTT | AAG����5��
| ��
������ֱ���߱�ʾ���ĶԳ��ᣬ�����ࡰ����������˳����GAATTC��CTTAAG������ǻ���˳��palindrome����ʵ��ͷ��ʾ��˫���Ͻ����и��λ�ã��и������5������G��AATTC����3����3������CTTAA��G����5������DNAƬ�Σ�����һ������ĩ�ˣ����������ǻ����ģ���ͨ���γ��������ճ�ϡ�����һ������ͬһλ�����и�˫��������ƽͷĩ�ˡ�����Hae���ʶ��λ���ǣ�
����������������������������������������
5������GG��CC����3��
3������CC��GG������
��
�����ڼ�ͷ��ָ���и����������DNAƬ���ǣ�
����5������GG CC����3��
������
����3������CC GG����5��
������ʱ����������������ø��ʶ�������˳����и�λ�ö���ͬ�����ֻ���ڵ�ʶ��˳�����м����ĺ�����ʱ��һ������������ø�����и��һ�����ܡ�����Hpa���Msp���ʶ��˳����5������GCGG����3�������������5��-������ण���ֻ��Hpa���ܹ��и��Щ����ͬ�е��ø��Ϊͬ��ø����Դͬ��ø��isoschizomer����
��������������������øҲ��רһ��ʶ��˳�����ǶԳƵĻ���˳������ʶ��˳���Ա���������ԵĹ̶�λ�����и�˫�������⼸�����������������ġ���ˣ���������������ø�и�������һ������DNAƬ�Σ����и��ֵ���ĩ�ˡ�����ڿ�¡������¡DNAƬ��û�ж���ô���
������������ø
����ϸ��������-����ϵͳ�е������������ɼ���ø��methylase������ɵġ�����øͬ����������ø������ȫ��ͬ��ʶ��˳����øʹʶ��˳���е�ij�������������������DNA��������������ø�п���
�������������Ŀǰֻ����5-������ण�M5C����M5Cռ����������е�2��7%����Ӭ��ijЩ�������⣩��M5C�����M5CpG����ʽ���ڣ���ͬ���ֻ�ͬһ���ֵIJ�ͬ��֯�У�M5C���ֵ�Ƶ��Ҳ������ͬ��Ŀǰ�Ѿ�������Ϊ��Ʒ���۵ļ���ø����23-2��
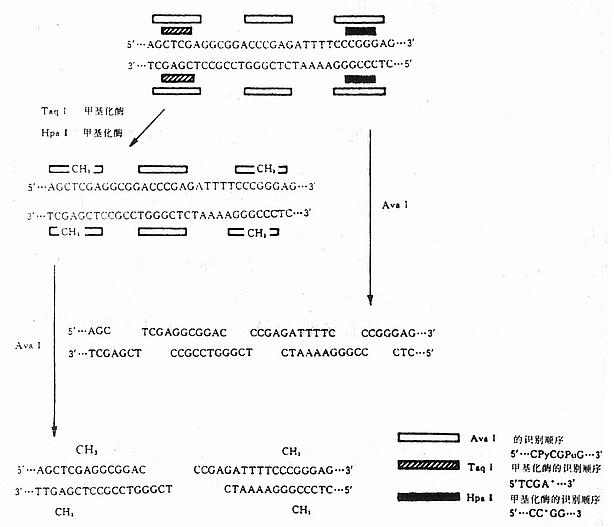
����������������ø�ͼ���øͬʱ���ã���ʹ�м��ֿ���ʶ��˳�������������øֻ������һ��ʶ��˳����Ч����������������øAva���ʶ��˳���ǣ�
5������CPyCGPuG����3��
3������GPuGCPyC����5��
����Py�������κ�һ����ण�pu�������κ�һ�����ʡ���˿�����4��ʶ��˳�����ͬʱʹ�ü���øTaq��ͼ���øHpa����Ava���ʶ��˳��ֻ��5������CCCGAG����3��������ͼ��˵����ͼ23-1����
���������һ��DNAƬ�Σ���Ava���3��ʶ��˳����Ava������ɵ�4��Ƭ�Ρ����ǣ��������Taq�����ø��Hpa�����øʹDNA˳���е���Щ���������Ȼ������Ava��ȥ�У����ֻ��1��ʶ��˳��ɱ�Ava�����ã�ֻ����2��DNAƬ�Ρ�
ͼ23-1 ��������������ø�ͼ����ļ��
��23-2������ø�������ļ��
����ø���� |
�����ļ�� |
| Alu�� | 5������AGC*T����3�� |
| BamH�� | 5������GGATC*C����3�� |
| Cla�� | 5������ATCGA*T����3�� |
| dam�� | 5������GA*TC����3�� |
| EcoR�� | 5������GAA*TTC����3�� |
| Hae�� | 5������GCC*C����3�� |
| Hha�� | 5������GC*GC����3�� |
| Hpa�� | 5������CC*GC����3�� |
| Hph�� | 5������TC*ACC����3�� |
| Msp�� | 5������C*CGG����3�� |
| Pst�� | 5������CTGCA*C����3�� |
| Taq�� | 5������TCGA*����3�� |
������*��ʾ����
�����������ӿ�¡������һЩø
������һ��DNA�ۺ�ø
������DNa polymerase���������ǽ�1�����������������ӵ����primer����3��-OH�ϣ��ͷų�һ����������ӣ�ppi��������ʽ��ʾ��
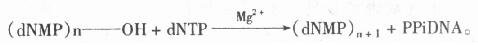
�����ۺ�ø��Ҫ�����¼��֣�
����1���˾�DNA�ۺ�ø���˾�DNA�ۺ�ø��E.Coli DNA polymerase����Ҫ��3�����ã���5����3���ľۺ����á������Ǹ���Ⱦɫ�������DNA���DNA�ϵĿ�϶�����г�RNA��������µĿ�϶����3����5��������ø���ԡ������ھۺ������в���Ĵ�������ᡣ��5����3������ø���ԡ��г������˵�DNA�������п�ƽ�ƣ�nick translation���е�Ӧ�ã�����������ܡ�
�����˾�DNA�ۺ�ø���KlenowƬ����������DNA�ۺ�ø���һ��Ƭ�Σ�ֻ����5����3�ۺ�ø���Ժ�3����5������ø���ԣ�ʧȥ��5����3����ø���ԡ����������DNA����ĩ�˳�Ϊ˫�����������32P��ǵ�����������ᣬ���ʹDNA����ͬλ�ر�ǡ����ý����и������ø�гɴ��е���ճ��ĩ�˵�DNAƬ�Σ�Ҫ�ñ��г�ƽͷĩ�˵�DNAƬ������ʱ����������KlenowƬ��ʹճ��ĩ�˵ĵ��������Ϊƽͷ��Ȼ����DNA����ø�����°�����DNAƬ������������
��������д˾�DNA�ۺ�ø��ʹ˾�DNA�ۺ�ø��ǰ�߲������õ���DNA��poly(dA-dT)Ϊģ�塣����þ���Ӻ�dNTPʱ���ܱ��ֳ�ø���ԣ����Է��ϳ�DNA�����á�����û��5����3����ø���ԣ�Ҳ�����õ���DNA��Ϊģ�塣
����2���ɾ���DNA�ۺ�ø����������T4DNA�ۺ�øΪ������Ҳ����5����3�ۺ�ø���ԣ�����������ø���Աȴ˾���Ҫ��200������ˣ���Ҳ���������뵥��ĩ�˻���ͬλ�ء�
����������RNA�ۺ�ø
����RNA�ۺ�ø��RNa polymerase����������ת¼RNA���е�RNA�ۺ�ø�бȽϸ��ӵ��ǻ��ṹ����˾�RNA�ۺ�ø������������������һ���ٽ���RNA���ӺϳɵĦ����ӣ����������ɵ��Ǧ�2�¦¦ҡ����ֽṹ��Ϊȫø��holoenzyme��,��ȥ�˦����ӵ�ø��Ϊ����ø���ɾ���RNA�ۺ�ø��û���ǻ���
������������RNA�ۺ�ø�����ࡣRNA�ۺ�ø������ں����У�ת¼rRNA˳��RNA�ۺ�ø������ں����У�ת¼�����������Ҫ��TATA����RNA�ۺ�ø������ں����У�ת¼���ټ��ֻ�����tRNA������5SrRNA������Щ�ظ�˳����Alu˳�����Ҳ������øת¼�������ᵽ�ġ�TATA�����ֳ�Goldberg �CHogness˳����RNA�ۺ�ø��ĽӴ��㣬������ø��ת¼��λ�����еġ�������������ת¼�����5����һ�࣬��ת¼�������20��30��������֮����һ�θ���AT��˳������ת¼��ʼ��Ϊ0������-33��27����������-27��21������֮�䣬��һ����TATA����һ����7�������ᡣRNA�ۺ�ø��ת¼��λ�ĵ��ͽṹ��ͼ23-2��ԭ��������Ҳ���ơ�TATA����Ľṹ��RNA�ۺ�ø�����ڡ�TATAAT����Pribnow���к͡�TTGA��CA������
ͼ23-2 ��ת¼ø�����û���
������������ת¼ø
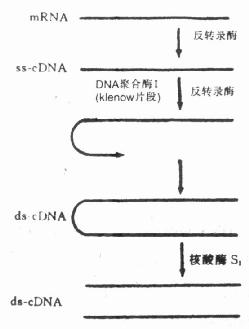
������ת¼ø��Reverse transcripatase������RNAΪģ��ָ������������������ϳɻ���DNA��cDNA����ø��������C�Ͳ����ķ�ת¼ø������B�Ͳ����ķ�ת¼ø����һ��������������RNA�����ķ�ת¼ø���������ǻ��ṹ�����������Ҳ����������в�ͬ�ṹ�ķ�ת¼ø������ø��Ҫþ���ӻ���������Ϊ�������ӣ�����mRNAΪģ��ʱ���Ⱥϳɵ���DNA��ssDNA�������ڷ�ת¼ø��DNA�ۺ�ø�������£��Ե���DNAΪģ��ϳɡ����С��͵�˫��DNA��dsDNA�������ɺ���øS1�гɶ���������˫��DNA����ˣ���ת¼ø���������κλ����mRNA��ת¼��cDNA������Ȼ��ɴ�����������������cDNA��Ҳ���������cDNA��Ϊ�����Եķ���̽�롣
�������ģ���������ø
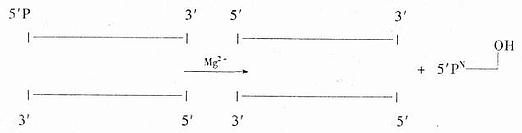
�����˾���������ø��exonuclease ���ǴӴ�3��-OHĩ�˵�˫��DNA����3����5�������г�5���������
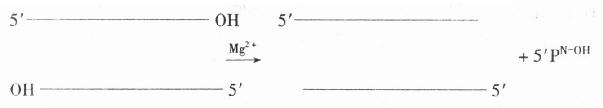
�����˾���������ø�����Ǵӵ���DNA��3���˺�5�����г��Ѻ����ᣨ������ʽ����
������1����2��
����ǰһ��ø��Ҫþ�������������Ӳ��л��ԣ���һ��ø����þ���ӣ���˼�ʹ�������ϼ�EDTA�������Ҳ�л��ԡ���һ�ִ��ɾ���˸�Ⱦ�Ĵ˾�����ȡ�Ħ˺�������ø�ǴӴ���5������ĩ�˵�˫��DNA�ϣ�����г�5���������ᣬ�ڷ�Ӧʱ��Ҫþ���ӣ�
�������壩����øS1
��������øS1��nuclease S1����Ҫ�ǽ��ⵥ��DNA����RNA���Ե���DNA�Ļ��Ը��ߡ�������;���г�DNAƬ�εĵ���ĩ��ʹ֮��Ϊƽͷĩ�ˣ��п��ϳ�dscDNAʱ�γɵġ����С����Լ�����DNA-RNA�Ӻ��ӵĽṹ��
����������DNAø
����DNAø��DNase�������ˮ��˫������DNA��һ������ø��ʹDNA���ӽ���ɴ���5������ĩ�˵ĵ�������Ѻ�����Ļ���
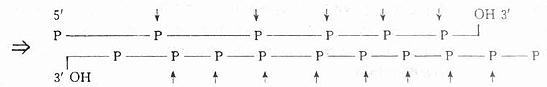
�����ڽ�������DNA�о�ʱ������ע���ֹDNAø����Ⱦ�������Ʊ���DNA��Ʒ���ή�⡣���������Լ����´�������Ʒ�м�EDTA������ñ������ƻ�������ø���ԡ�
�������ߣ�RNAø
����RNAøA��ribonuclease A����������ऺ������3��������ϣ��п������ں��������ӵ�5�����������һ��RNAøT1ֻ�����������ʺ������3����������п������ں��������ӵ�5�������������ķ���������Һ����Һ�ж�����RNAø������ڲ���RNA��Ʒʱ����������ף�ʵ���õIJ�������Ҫ��250��濾4h��RNAø���ȣ�������RNAø�����Ƽ�������
�������ˣ�����ø
������õ���T4DNA����ø��T��DNa ligase������˫��DNA�����ڵ�3��OH��5�������֮���γ�����������������������Ӿ���ճ��ĩ�˵�����DNAƬ�Σ�����������ƽͷĩ�˵�DNAƬ�Σ�ʹ֮��Ϊһ������DNA���ӡ���������øֻ������˫��DNA���������ӵ���DNA���ӡ�T4RNA����ø����ڵ���DNA���ӻ�RNA���ӵ�5���������3��-OH֮������ɹ��ۼ���
�������ţ�ĩ��ת��ø
����ĩ������������ת��ø��terminal deoxynucleotide transferase���������ǽ�����������ӵ�DNA���ӵ�3��-OHĩ�ˡ�
������ʮ����������ø
������DNA��RNA���������������Ǻ��������������Ǻ������ϳ�5��-������л���һ�����;��������ø����DNA��RNA����5�����ϱ��32P���ھ�����������øø��DNAƬ�κ��ü�������ø��alkaline phosphatase������������ֹø�е�Ƭ���������ӣ����ڿ�¡DNAƬ��ʱ�ر����á�