�ھ��¡����ʴ�л����
(Regulation in Metabolism)
�������ʴ�л����������Ļ���������������������ʻ������������ʴ�л�������������ĺ���صĴ�л;������ɣ�����л;��(���ǵ�������֬����ĺϳɵ�)������һϵ�е�ø�ٻ�ѧ��Ӧ��ɡ�����������£����ִ�л;������ȫ�����������������н��ࡢ�й��ɵؽ��У�ͬʱ��Ϊ��Ӧ��������ı仯����ʱ�ص�����Ӧ�ٶȣ���������Ķ�̬ƽ�⡣�ɼ����������ʴ�л�������ܵĵ����½��еġ�
������л���ڻ����ձ����������磬�������ڳ��ڽ������������γɵ�һ����Ӧ�����������̶�Խ�ߵ�������л���ڵĻ���Խ���ӡ���ϸ����������ϸ���ڴ�л��Ũ�ȱ仯��Ӱ�죬�ı���������ø�Ļ��Ժ�ø�ĺ������Ӷ����ڴ�л���ٶȣ�����ϸ��ˮƽ�Ĵ�л���ڣ����������ڽ����Ͻ�Ϊԭʼ�ĵ��ڷ�ʽ���ϸ��ӵĶ�ϸ������������ڷ���ϸ�����ߵȶ����������ר�ŵ��ڷ������٣���Щ���������ڵļ��ؿ��Զ�����ϸ�����Ӵ�л�������á����ؿ��Ըı�ijЩø�Ĵ����Ի�����Ҳ���Ըı�ϸ���ڴ�л���Ũ�ȣ��Ӷ�Ӱ���л��Ӧ���ٶȣ����Ϊ����ˮƽ�ĵ��ڡ��ߵȶ��ﲻ�����������ڷ���ϵͳ�����һ��й��ܸ��ӵ���ϵͳ���������Ŀ����£�����ͨ�����ʶ�ЧӦ��ֱ�ӷ���Ӱ�죬����ͨ���ı�ijЩ���صķ��ڣ�������ijЩϸ���Ĺ���״̬����ͨ�����ּ��صĻ���Э�����������л�����ۺϵ��ڣ����ֵ��ڼ�������ˮƽ�ĵ��ڡ�����������ϸ��ˮƽ�Ĵ�л���ڡ�����ˮƽ�ĵ��ں�����ˮƽ�ĵ��ڣ��ڸߵȶ����������ȫ�����ڣ�����ֱ���н��ܡ�
��һ�ڡ�ϸ��ˮƽ�Ĵ�л����
����һ��ϸ����ø�ķָ��ֲ�
���������ʴ�л�����п�֪��ø��ϸ�����Ƿָ��ŷֲ��ġ���л���йص�ø���������һ��ø��ϵ���ֲ���ϸ����ijһ����У�����,�ǽͽ�øϵ����Ԫ�ϳɡ��ֽ�øϵ�����ڰ�Һ�У�������ѭ��øϵ��֬�����-����øϵ��λ�������壻����ϳɵ�øϵ����ּ�����ϸ�����ڡ�������ø�ĸ���ֲ�Ϊ��л���ڴ���������������ʹijЩ�������ؿ��Խ�Ϊרһ��Ӱ��ijһϸ������е�ø�Ļ��ԣ�������Ӱ����������е�ø�Ļ��ԣ��Ӷ���֤�����巴Ӧ�������ԡ�һЩ��л��������ڸ�ϸ����ּ�Ĵ����ƶ�Ҳ���Ըı�ϸ����ijЩ��ֵĴ�л�ٶȡ����磬�ڰ�Һ�����ɵ�֬����øA��Ҫ���ںϳ�֬���������ⶾ��������£����ⶾ��֬��ת��ø�Ĵ���֬����øA�ɽ��������壬�����-�����Ĺ��̡����磬Ca�����Ӽ�ϸ���������г���,���Դٽ���Һ�е���Ԫ�ֽ⣬��Ca������������������������Ԫ�ϳɡ�
�������ʴ�лʵ������һϵ�е�ø�ٷ�Ӧ����л�ٶȵĸı䲢�������ڴ�л;����ȫ��ø���Եĸı䣬������ֻȡ����ijЩ����ijһ���ؼ�ø���Եı仯����øͨ��������ͨ·�д�����һ����Ӧ��ø����Ϊ����ø�����Ļ��Ըı䲻������Ӱ������ø��ϵ����Ӧ�����ٶȣ����������Ըı��л��Ӧ�ķ����磬ϸ����ATP/AMP�ı�ֵ���ӣ���������������Ǽ�ø(�ͱ�ͪ�ἤø)�Ļ��ԣ��ⲻ���������ǽͽ���ٶȣ�������ͨ���������-1��6-������ø��ʹ�Ǵ�л��������������������ˣ��ı�ijЩ�ؼ�ø�Ļ��������ڴ�л���ڵ�һ����Ҫ��ʽ��
���������л��ϸ��ˮƽ���ڣ����ٶȷ�����˵�����ַ�ʽ��һ���ǿ��ٵ��ڣ�һ����������������ڼ��ɷ��������ֵ�����ͨ���������������ԭ�е�ø����������ø�ٷ�Ӧ�ٶȵģ������¶ȡ�pH���������ø�����ز��������£�ͨ���ı�ø���ӵĹ�����ø���ӽ��л�ѧ������ʵ��ø�ٷ�Ӧ�ٶȵ�Ѹ�ٸı�ġ���һ���dzٻ����ڣ�һ�㾭��Сʱ�����ʵ�֡����ַ�ʽ��Ҫ��ͨ���ı�ø���ӵĺϳɻ��ٶ�������ϸ����ø���ӵĺ������ֽ�������������÷������£�
��������ø���ӽṹ�ĵ���
����(һ)�乹����
����1.�乹���ڵĸ��ijЩ��������ø�����ϵķǴ���λ����ؽ�ϣ�����ø���ķ��ӹ������ı䣬�Ӷ��ı�ø�Ļ��ԣ����������Ϊø�ı乹���ڻ�Ʊ�λ����(allosteric regulation)�������ֵ������õ�ø��Ϊ��ø��乹ø(allosteric enzyme)����ʹø�����乹ЧӦ�����ʳ�Ϊ�乹ЧӦ��(allosteric effector);��乹������ø���Ե���ǿ�����ЧӦ����Ϊ����乹��(allosteric activator)����ЧӦ���֮���Ϊ���Ʊ乹��(allosteric inhibitor)��ЧӦ��乹������������ձ���ڣ����������ڿ��ٵ���ø���Ե�һ����Ҫ��ʽ���ֽ�ijЩ��л;���ı乹ЧӦ���б����£���
��9-1���Ǻ�֬����лøϵ��ijЩ�乹ø����乹ЧӦ��
| ��л;�� | �乹ø | ����乹�� | ���Ʊ乹�� |
| �������ֽ� | ���Ǽ�ø | �� | G-6-P |
| �� | ������Ǽ�ø | AMP��ADP��FDP��Pi | ATP�������� |
| �� | ��ͪ�ἤø | FDP | ATP������CoA |
| �� | ������������ø | AMP | ATP������֬��CoA |
| �� | ������ϳ�ø | ADP��AMP | ATP |
| ������ | ����-1��6-������ø | �� | AMP |
| �� | ��ͪ���ǻ�ø | ����CoA��ATP | �� |
| ֬����ϳ� | ����CoA�ǻ�ø | �����ᡢ�������� | ����֬��CoA |
����2.�乹���ڵ��������塡�乹ЧӦ��ø�Ŀ��ٵ�����ռ���ر���Ҫ�ĵ�λ����ǰ���Ѿ��ἰ����л�ٶȵĸı䣬����������Ӱ����������лͨ·�д���һ����Ӧ��ø��������л��Ӧ������ø�Ļ��Զ�����ġ���Щø�Ե��ﲻ���������϶���ѧԭ�����������ܵ�һЩ��л������ƻ���Щ���ƻ����ô����ͨ���乹ЧӦ��ʵ�ֵġ����,��Щø�Ļ������Լ��������ܵ���л����Ũ�ȵĵ��ڣ���Ի����������л���ؾ�����Ҫ�����塣���磬�乹ø��������������л�ĵ��ھ�����Ҫ���塣����Ϣ״̬�£������������Ľ��ͣ�ATP��ϸ���ڻ��ۣ���ATP��������Ǽ�ø�����Ʊ乹�������Ե���F��6��P��G��6��P�Ļ��ۣ�G��6��P�������Ǽ�ø�����Ʊ乹�����Ӷ����������ǵ������ֽ⡣ͬʱ��ATPҲ�DZ�ͪ�ἤø��������ϳ�ø�����Ʊ乹��������ǿ�˶������������ֽ�����ƣ��Ӷ�������ATP�Ľ�һ�����ɡ���֮��������ATP���ٶ�ADP��AMP����ʱ��AMP������ƹ��Ǫ�1��6��������ø��������������ͬʱ����������Ǽ�ø��������ϳ�ø��ø�������ǵķֽ���������������ATP�����ɡ�������ͨ���乹���ڣ�ʹ����ATP�����ɲ��¹������٣���֤�˻������Դ����Ч���á�
ͼ9��1���乹ø�ĵ���Ũ������
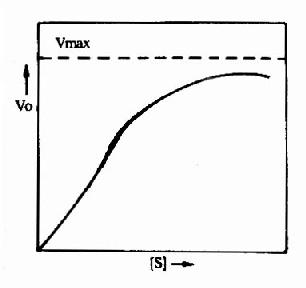
����3.�乹���ڵĻ�����Ŀǰ��֪�����ܱ乹���ڵ�ø�������������������ǻ���ɵľۺ��塣�е��ǻ����������ϣ�������ã���Ϊ���ǻ����е��ǻ���乹����ϣ����ӵ������ã��Ƶ����ǻ�����Ҳ����ͬһ�ǻ��ϼȴ��ڴ���λ�ִ��ڵ��ڲ�λ���乹��������ǻ�(��λ)���Ƿǹ��ۼ��Ľ�ϣ���Ϻ�ı�ø�Ĺ���(��,��Ϊ���ɻ����)���Ӷ�ʹø���Ա����ƻ�乹ø���ף�����ø��ͬ���䶯��ѧ�����������Ϸ���ʽ��ø�ٷ�Ӧ�ٶȺ�������Ũ�ȵĹ�ϵ���߲��ʾ��ζ�������S�Σ�S������������Ѫ�쵰�Ľ�����������(ͼ9��1)��
�������乹��������ǻ�(��λ)��Ϻ������ø���ӵĹ�����ʲô����Ӱ����?�����Թ�-1��6-������øΪ��������һ���̡�����-1��6-������ø�����ĸ��ṹ��ͬ���ǻ�����ɣ�ÿ���ǻ��ķ�����ԼΪ310��000Da��ÿ���ǻ��ϼ��д���λҲ�е��ڲ�λ���ڴ���λ���ܽ��һ����FDP���ڵ��ڲ�λ���ܽ��һ���ӱ乹������ø�����ִ�����ʽ����������(T�͡�����)���ɳ���(R�͡��ͻ���)��AMP�Ǵ�ø�����Ʊ乹������ø����T��ʱ��������ڲ�λת���ۺ����ڲ���������AMP��ϣ��ʶ�AMP�����ж����ֳ��ϸߵĻ��ԡ��ڵ�һ��AMP��������ڲ�λ��Ϻ�T����ת���R�ͣ����ǻ�������̷����ı䣬���ڲ�λ��̱�¶����AMP�����������ӣ�ø�Ļ�������������ǹ���-1��6-������ø�ɽ����ͱ���ɳ��͵ı乹���̡����Ʊ乹���ٽ����������ͻ����͵�ת�䣬����乹����ٽ��ͻ������������͵�ת�䡣��һ�乹�����ǿ����(ͼ9��2)��ͼ��3���������ȩ��֬���ᣭ���嵰��ʹ������ת��Ϊ�����͡�
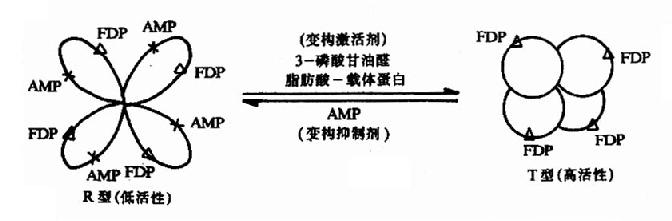
ͼ9��2�����ǣ�1��6������ø�ı乹ЧӦ��
����ø�ǻ��ϵĴ���λ��X��ø�ǻ��ϵĵ��ڲ�λ��FDP������-1��6��������
�����乹ЧӦ��������ø�ĵ��Ҳ������øϵ���ղ�����е��������ǽṹ��ͬ�����������һ��˵������С�������ʡ�һ��ø���ж��ֱ乹ЧӦ�����ڡ�
�������ǣ�1��6��������ø�ı乹������T����R�͵Ŀ���ת�䡣��Щø�ı乹ЧӦ���ɱ���Ϊø���ӵľۺϻ��ۣ�������CoA�Ȼ�ø������֬����ϳɹ����еĹؼ�ø�����������ֲ�ͬ�ǻ����ɵ�ԭ���壬ÿ���ǻ��в�ͬ�Ĺ��ܣ��ֱ��ǣ����������嵰�ף����ܽ�ϸ��������أ��������Ȼ�ø�����ܴ������ط����Ȼ���Ӧ���Ȼ�ת��ø�����ܽ��������ϵ��Ȼ�ת�Ƹ�����CoA�γɱ�������CoA���͵����ǻ�������������������������ϣ�ʹԭ����ۺ�Ϊ����塣Kieinschmidt�����ڵ��������¿����������������������ʹԭ����ۺ��γɵ���ά״�Ķ����(ͼ9��3)��ֻ�ж����ø���д����ԡ�ATP��Mg������ʹ�������Ϊԭ�����ʹøʧ�����֬����CoA����������Ĵپۺ����ã���ˣ����Ƕ��Ǹ�ø�ı乹���Ƽ���
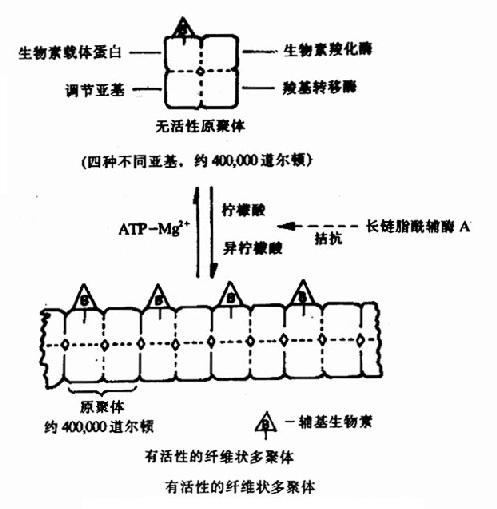
ͼ9-3������CoA�Ȼ�ø�ۺϽ��ʾ��ͼ
����(��)ø���ӻ�ѧ���ε���
����1.ø���ӻ�ѧ���εĸ���
����ø���������ϵ�ijЩ���ſ�����һ��ø�Ĵ��·�������Ĺ������Σ��Ӷ�����ø���Եĸı䣬������̳�Ϊø��ø�ٻ�ѧ����(chemical modification)�������ữ�������ᣬ��������ȥ�����������ջ���ȥ���ջ���������ȥ�����Լ�-SH���ͣ�S��S��������ȣ��������ữ�����������������ʴ�л��������Ϊ������
����ϸ���ڴ����Ŷ��ֵ���ø(Protein Kinase)�����ǿ��Խ�ATP�����еĦ�-�������ת�����ض��ĵ����ӵ����ϣ�ʹ�������ữ(phosphorylation)�����ữ��Ӧ���Է�����˿���ᡢ�հ�����Ұ���л��ϡ���˿������հ���л����ữ��øͳ��Ϊ����˿����/�հ��ἤø(Protein Serine/Threonine Kinase)�����Ұ���л����ữ��øͳ��Ϊ�����Ұ��ἤø(ProteinTyrosine Kinase)��������Ӧ�ģ�ϸ����������Ŷ��ֵ���˿����/�հ�������ø(ProteinSerine/Threonine Phosphotase)�͵����Ұ�������ø(Protein Tyrosine Phosphotase)�����ǿɽ���Ӧ�����������ȥ��ø�Ļ�ѧ������乹����һ����Ҳ�ǻ������ʴ�л�п��ٵ��ڵ�һ����Ҫ��ʽ����9?�г���һЩø��ø�ٻ�ѧ���ε�ʵ����
����9��2��ijЩø��ø�ٻ�ѧ���ε���
| ø�� | ��Ӧ���� | ЧӦ |
| �������ữø | ���ữ�������� | ����/���� |
| ���ữøb��ø | ���ữ�������� | ����/���� |
| ���ữø����ø | ���ữ�������� | ����/���� |
| ��Ԫ�ϳ�ø | ���ữ�������� | ����/���� |
| ��ͪ������ø | ���ữ�������� | ����/���� |
| ֬��ø��֬��ϸ���� | ���ữ�������� | ����/���� |
| �Ȱ������ϳ�ø���˾��� | ���ջ��������� | ����/���� |
| ���������������⣩ø | ��SH/-S-S- | ����/���� |
����2.ø�ٻ�ѧ���εĻ���
����������Ԫ���ữø��ø�ٻ�ѧ�������о��ñȽ������һ�����ӡ���ø��������ʽ�������Ե����ữøb���л��Ե����ữøa�����ữøb�Ƕ����壬������ԼΪ85��000Da������ø�Ĵ��£�ʹÿ���ǻ��ֱ����ATP������һ��������ţ�ת��Ϊ���ữøa�����߾��и��ԡ����������ữøa����������پۺϳɻ��Խϵ͵�(���ڸ��ԵĶ�����)���ữøa�ľ���(ͼ9��4)��
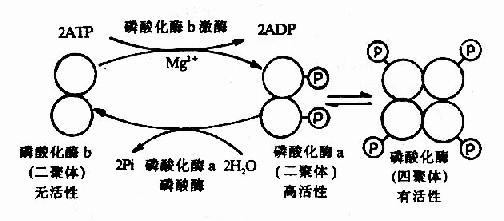
ͼ9��4���������ữø��ø�ٻ�ѧ��������
����3.ø�ٻ�ѧ���ε��ص�
����(1)�������ø�ٻ�ѧ���ε�ø����������(��ͻ���)���л���(�����)������ʽ������֮��Ļ��䷴Ӧ,���������й��۱仯���ɲ�ͬ��ø���д��������⻥�䷴Ӧ��ø���ܻ����������(�缤��)�Ŀ��ơ�
����(2)�����ٲ�ʽЧӦ������ø�ٻ�ѧ������ø�����ķ�Ӧ�������ٲ�ʽ(�Ŵ�)ЧӦ�������ĵ������ؾͿ�ͨ����������ø�ٷ�Ӧ��ʹ��������һ��ø������ѧ���Ρ���ˣ����෴Ӧ�Ĵ�Ч�ʳ��ϱ乹����Ϊ�ߡ�
����(3)���ữ���������dz�����ø�ٻ�ѧ���η�Ӧ��һ�����ǻ��������ữ��������һ����ATP������ϳ�ø���������ĵ�ATP��ȣ���Ȼ���ٵöࣻͬʱø�ٻ�ѧ�������зŴ�ЧӦ����ˣ����ֵ��ڷ�ʽ��Ϊ������Ч��
����(4)���ֵ���ͬ�乹����һ��,��������������Ҫ�����С���ǰ���ļ�����Ԫ���ữø�Ļ�ѧ���ι����У���ϸ��Ҫ������ֹͣ��Ԫ�ֽ�,�����ữøa�����ữøa����ø�Ĵ��¼�ˮ����ȥ�������ת������Ե����ữøb���Ӷ�������ֹͣ����Ԫ�ķֽ⡣
�������⣬ø�ٻ�ѧ������乹����ֻ��������Ҫ�ĵ��ڷ�ʽ����ijһ��ø��˵��������ͬʱ�������ַ�ʽ�ĵ��ڡ��磬��Ԫ���ữø�ܻ�ѧ���ε�ͬʱҲ��һ�ֱ乹ø����������ÿ���ǻ����д���λ�͵��ڲ�λ��������AMP�������ATP���ƣ������ڱ乹���ڡ�ϸ����ͬһ��ø��˫�ص��ڵ�����������ڣ��乹������ϸ����һ�ֻ������ڻ��ƣ�������ά�ִ�л�������ƽ�������Ҫ���ã�����ЧӦ��Ũ�ȹ��ͣ���������ȫ��ø���ӵĵ��ڲ�λ���ʱ���Ͳ��ܶ�Ա���е�ø�������ã�������Ӧ��������Ӧ��������£������������������ͷţ�����ͨ��cAMP,����һϵ�е��ٲ�ʽ��ø�ٻ�ѧ���η�Ӧ������ת�����ữøb��Ϊ�л��Ե����ữøa��������Ԫ�ķֽ⣬Ѹ����Ч���������ļ��衣
��������ø��������
������ͨ���ı�ø���ӵĽṹ������ϸ����ԭ��ø�Ļ����⣬�����廹��ͨ���ı�ø�ĺϳɻ��ٶ��Կ���ø�ľ��Ժ��������ڴ�л��Ҫ����ij��ø��Ũ�ȣ�������ø���ϳɵ��յ�����������⣬������ͬʱ����ø������ٶȣ��ַ������£�
����(һ)ø���ϳɵ��յ������
����ø�ĵ�����������Լ�ҩ��ȶ�����Ӱ��ø�ĺϳɡ�һ�㽫��ǿø�ϳɵĻ������Ϊ�յ���(inducer)������ø�ϳɵĻ������Ϊ�����(repressor)���յ��������������ת¼ˮƽ����ˮƽӰ�쵰���ʵĺϳɣ�����Ӱ��ת¼���̽�Ϊ���������ֵ�������Ҫͨ��һϵ�е���������ϳɵĻ��ڣ��ʵ���ЧӦ���ֽϳٻ�����һ��ø���յ��ϳɣ���ʹ��ȥ�յ�����ø���ܱ��ֻ��ԣ�ֱ��ø��������ϡ���ˣ����ֵ��ڵ�ЧӦ����ʱ��ϳ���
����1.�����ø�ϳɵ��յ����á���ø���ĵ��ﳣ�������յ���ø�ĺϳɣ���������������ձ���ڡ��ߵȶ������ڣ����м��صĵ������ã������յ����ò�������������Ҫ������,ijЩ��л;���еĹؼ�øҲ�ܵ�����յ����ڡ����磬������������ҵ�������8%����70%��������еľ�����ø�Ļ��Կ�����2?������ʳ���������պ�Ѫ�ж��ְ������Ũ�����ӣ�������Ũ�ȵ������ֿ����յ�������ֽ�ø��ϵ�еĹؼ�ø�����հ�����ˮø���Ұ���ת��ø��ø�ĺϳɡ������յ����ö���ά���������백����Ũ�ȵ���Ժ㶨��һ�����������塣
����2.�����ø�ϳɵ��������л��Ӧ���ղ��ﲻ����ͨ���乹����ֱ������ø��ϵ�еĹؼ�ø�������ʼ��Ӧ���õ�ø����ʱ���������Щø�ĺϳɡ����磬�ڵ��̴�������ϳ��У��£��ǣ��£����������øA(HMg CoA)��ԭø�ǹؼ�ø�����ܵ��̴��ķ�������������ַ������ֻ�ڸ�������з�������ճĤ�е��̴��ĺϳ��ƺ��������ַ������ڵ�Ӱ�졣�����ʳ�������̴��������̴��������ߵ�Σ�ա����⣬���-����-��-ͪ����(ALA)�ϳ�ø������Ѫ���غϳ�øϵ�е���ʼ��Ӧø������Ѫ���صķ��������
����3.���ض�ø�ϳɵ��յ����á������Ǹߵȶ�������Ӱ��ø�ϳɵ�����Ҫ�ĵ������ء���Ƥ�ʼ������յ�һЩ������ֽ��л�������ʼ��Ӧ���õ�ø��������;���ؼ�ø�ĺϳɣ����ȵ��������յ��ǽͽ��֬����ϳ�;���еĹؼ�ø�ĺϳɡ�
����4.ҩ���ø�ϳɵ��յ�����
�����ܶ�ҩ��Ͷ���ɴٽ���ϸ�������е�����ø(��ƻ�Ϲ�������ø)������һЩҩ���лø���յ��ϳɣ��Ӷ��ٽ�ҩ�ﱾ��������ҩ�������ʧ���Է�ֹҩ�������ж����ۻ�������Ҫ�����塣�����õı��ʣ�Ҳ���ڵ����ø�ϳɵ��յ����á���һ����,��Ҳ����˶����³�����ҩ�����磬���ڷ��ñ��ͱ��IJ��ˣ����ͱ����յ����ɹ���ĵ�����ø��ʹ���ͱ���ҩЧ���͡����������������ʱ��Ҳ�����յ�Ҷ�ỹԭø�ĺϳɶ�ʹԭ�������İ�����ʲ��������ҩ��ʧЧ����
����(��)ø���ӽ���ĵ���
����ϸ����ø�ĺ���Ҳ��ͨ���ı�ø���ӵĽ����ٶ������ڡ���������£�������ø�Ļ������ӣ���Ҫ������ø��������ٶȼ������¡�����Ҳ��ʹ������øA�Ȼ�øŨ�Ƚ��ͣ��������ø���ϳɼ����й��⣬����ø���ӵĽ����ٶȼ�ǿ�йء����ͱ���ҩ���ʹϸ��ɫ��b5��NADPH��ϸ��ɫ�أ�450��ԭø������٣���Ҳ������ҩ��ʹ������ø������ǿ��һ��ԭ��
����ø������ϸ������ø���е���ˮ��ø�Ĵ������⣬���,���ܸı䵰��ˮ��ø���Ի�ˮ��ø����ø���ڷֲ������أ����ɼ�ӵ�Ӱ��ø���Ľ����ٶȡ��й�������˽ⲻ�ࡣ��֮��ͨ��ø�����Ե���ø��������Ҫ�Բ���ø���յ���������á�