参考资料
蛋白质一级结构的测定方法
研究蛋白质的一级结构从确定组成蛋白质的单元结构棗氨基酸算起,已有150年的悠久历史,直到1955年,Sanger首次阐明胰岛素的氨基酸排列顺序,为研究蛋白质的一级结构开辟了道路。这在分子生物学的发展进程中是一个重要突破。目前关于核酸的一级结构研究,由于Sanger等发明了加减法,可以得到了突飞猛进的发展。对此之下,关于蛋白质的一级结构研究进展不如核酸迅速。但随着Edman液相自动顺序分析仪和固相顺序分析仪以及气相色谱质谱(GCMS)等方法的相继出现。使结构分析的速度也显著加快。至今已完成近千种蛋白质的一级结构分析。目前不仅样品用量减少,而且工作人员也大大减少。当年Sanger分析胰岛素用了整整十年的时间,今天运用自动化仪器,分析一个分子量在10万左右的蛋白质只需要几天,可见新技术的应用和发展对科学发展起的促进作用,蛋白质一级结构测定方法的综述及专著文献较多,这里只扼要加以概述。
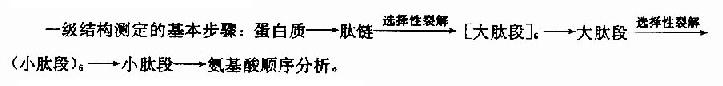
蛋白质分子的一级结构测定,概括起来包含多肽链的分离、降解、肽段的分离和顺序分析以及-S-S-定位等。
一级结构的测定方法可概述如下:
1.多肽链的分离
在测定一个蛋白质的结构以前,首先必须保证被测蛋白质的纯度,使结果准确可靠。其次要了解它的分子量和亚基数,按照其亚基数将蛋白质分成几个多肽链。
1)肽链的拆开
蛋白质分子多肽链的连接有共价结合和非共价结合两种。要拆开以共价结合的-S-S-连接的多肽链,必须采用的化学处理方法常有:
①过甲酸氧化
用氧化剂过甲酸断裂-S-S-。这个反应一般在0℃下进行2小时左右,两个S就全部能转变成磺酸基,这样被氧化的半胱氨酸称为磺基丙氨酸。
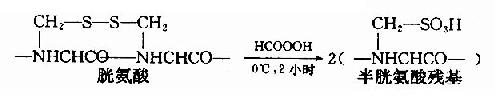
如果蛋白质分子中同时存在半胱胺酸,那么也会被氧化成磺基丙氨酸。此外甲硫氨酸和色氨酸也可被氧化,从而增加分析的复杂性。
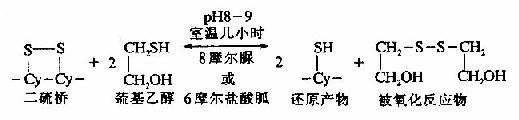
②巯基乙醇还原
利用还原剂巯基乙醇亦可使蛋白质的-S-S-断裂。当高浓度的巯基乙醇在pH8?条件下室温保温几小时后,可以使-S-S-定量还原为桽H。与此同时反应系统中还需要有8摩尔脲或6摩尔盐酸胍使蛋白质变性,多肽链松散成为无规则的构型,此时还原剂就可作用于-S-S-。此反应是可逆的,因此要使反应完全,疏基乙醇的浓度必需在0.1-0.5摩尔。
③Cleland试剂的还原作用
Cleland′s指出二硫赤苏糖醇(dithioerythriotol)及二硫苏糖醇(dithiothriotol)在氧化还原能力上是比较强的试剂,只要0.01摩尔就能使蛋白质的-S-S-还原,反应基本与疏基乙醇相似,且在许多球蛋白反应中,可以不用变性剂。
Cleland试剂首先与蛋白质-S-S-形成中间物,反应终了,还原剂被氧化形成一个稳定的六环化合物,蛋白质则被还原。

还原蛋白不稳定,SH基极易氧化重新生成-S-S-键。稳定SH基的方法有:
(A)烷基化试剂使SH基转变为稳定的硫醚衍生物。
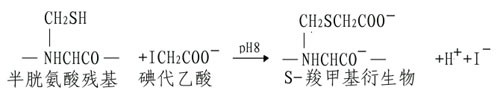
如果碘代乙酰胺代替碘代乙酸,其产物S羧氨甲基衍生物不带电荷,磺代乙酸也可与组氨酸、蛋氨酸和赖氨酸发生反应,但反应条件不同,可通过各种pH及反应时间进行控制。
(B)氨乙基化

蛋白质分子的几条肽链若以非共价健结合,则用尿素、盐酸胍等变性剂即可拆开。蛋白质的多肽链被拆开后,将它分离纯化,一般多用凝胶过滤、离子交换、电泳等方法,兹不赘述。
分离纯化后的每条肽链还要进一步分析其末端。
2)末端分析 其方法较多,这里我们只介绍较常用的几种。
(1)N-末端测定
A.二硝基氟苯法(FDNB,DNFB):1945年Sanger提出此方法,是他的重要贡献之一。
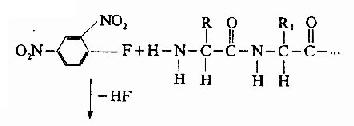

DNP-氨基酸用有机溶剂抽提后,通过层析位置可鉴定它是何种氨基酸。Sanger用此方法测定了胰岛素的N末端分别为甘氨酸及苯丙氨酸。
B.氰酸盐法:1963年Stank及Smyth介绍了一种测定N末端的新方法,步骤如下:
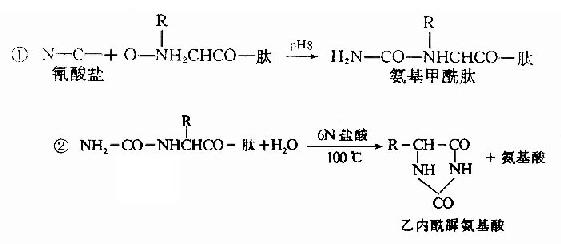
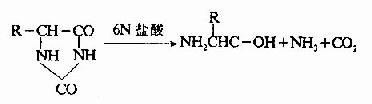
由于乙内酰脲氨基酸不带电荷,因此可用离子交换层析法将它与游离氨基酸分开,分离所得的乙内酰脲氨基酸再被盐酸水解,重新生成游离的氨基酸,鉴别此氨基酸即可了解N-末端是何种氨基酸。
C.二甲基氨基萘磺酰氯法:1956年Hartley等报告了一种测定N-末端的灵敏方法,采用1-二甲基氨基萘-5-磺酰氯,简称丹磺酰氯。它与游离氨基末端作用,方法类似于Sanger的DNFB法,产物是磺酰胺衍生物。
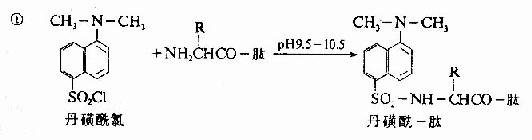
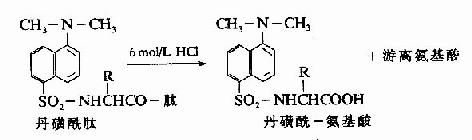
丹磺酰链酸具有强烈的黄色荧光。此法优点为灵敏性较高(比FDNB法提高100倍,样品量小于1毫微克分子)及丹磺酰氨基酸稳定性较高(对酸水解稳定性较DNP氨基酸高),可用纸电泳或聚酰胺薄膜层析鉴定。
(2)C-末端分析
A.肼解法:这是测定C-末端最常用的方法。将多肽溶于无水肼中,100℃下进行反应,结果羧基末端氨基酸以游离氨基酸状释放,而其余肽链部分与肼生成氨基酸肼。
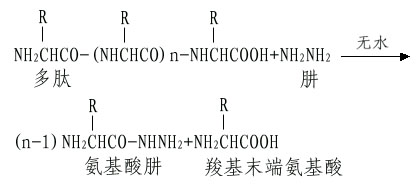
这样羧基末端氨基酸可以采用抽提或离子交换层析的方法将其分出而进行分析。如果羧基末端氨基酸侧链是带有酰胺如天冬酰胺和谷氨酰胺,则肼解时不能产生游离的羧基末端氨基酸。此外肼解时注意避免任何少量的水解,以免释出的氨基酸混淆末端分析。
B.羧肽酶水解法:羧肽酶可以专一性地水解羧基末端氨基酸。根据酶解的专一性不同,可区分为羧肽酶A、B和C。应用羧肽酶测定末端时,需要事先进行酶的动力学实验,以便选择合适的酶浓度及反应时间,使释放出的氨基酸主要是C末端氨基酸。
3)氨基酸组成分析
在进一步分析多肽链的氨基酸顺序之前,首先应了解它是由那几种氨基酸组成的,每种氨基酸有多少?分析组成的方法有:
①层析法
将多肽链完全酸水解成游离氨基酸,然后进行Dansyl标记,聚酰胺薄膜层析,此方法在蛋白质结构分析中是一种超微量的分析术,但此方法用于定量分析尚不够准确。
②离子交换层析法
Spaekman等发展了一种精确的氨基酸组分的定量方法。他们采用磺酸型的离子交换树脂,这是一种高分子量的固体聚苯乙烯,带有大量的功能基团,磺酸基在低pH和低离子强度条件下,根据氨基酸的酸碱性,氨基酸带正电,于是替换下树脂上的Na+,借助静电作用而结合到磺酸基上。
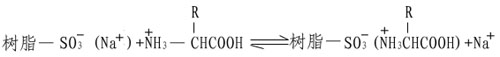
由于各种氨基酸在树脂上的亲和力不同,因此当改变溶液pH和离子强度,便可依次将它们洗脱下来而分开,并进行定量测定。在此基础上发展了氨基酸自动分析仪。随着科学技术的日益进展,氨基酸自动分析仪在样品的用量,分离速度及检测能力上也有了很大的提高。目前最好的仪器样品分析量只要几十Picomole,分析时间只要数十分钟,而且计算全部自动化,给研究蛋白质一级结构带来了极大的方便。
2.多肽链的降解
多肽链的氨基酸组成往往是比较复杂,因此直接分析多肽的氨基酸顺序还是很困难的,多采用将多肽链进一步降解成为更小的片段,然后再行分析。肽键的裂解是一级结构研究工作中的重要问题,它要求裂解点少,选择性强,而且反应产率高,目前主要有化学法和酶解法两类。
1)化学法
(1)溴化氰法 是最理想的化学方法,能选择性断裂甲硫氨酸所在的肽键
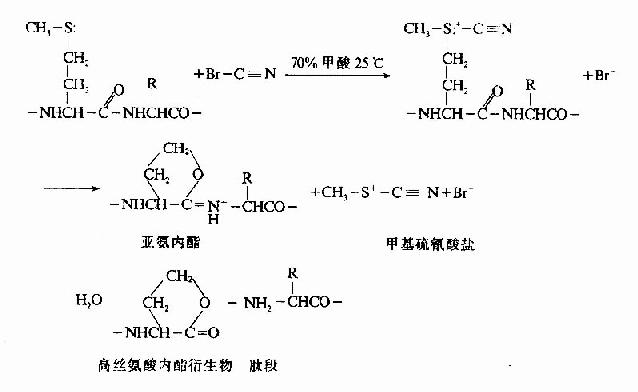
溴化氢化学降解法其优点:
①一般蛋白质含甲硫氨酸较少,由此可获得大片段
②专一性强
③产率高达80%以上
④作用条件温和,在室温中用几到十几小时即可。
(2)部分酸水解法
Sanger在分析胰岛素的一级结构中采用了此法,即用0.1N盐酸在110℃或用6N盐酸在37℃水解。这种部分酸水解的方法特异性不强,因此对大片段的蛋白质和肽均不合适。
(3)羟胺法
这种方法近十年来开始受人注意,羟胺能专一性地裂解AsnGly的肽键,酸性条件下裂解Asn-Pro肽键。已用于某些蛋白质的分析。
(4)N-溴代琥珀酰亚胺法
主要裂解Try处的肽键,五十年代研究较多。但由于它也能断裂TyrHis肽键,因此应用不广。
2)酶解法
酶水解法较化学法具有更多的优越性,使用也更广泛。因其具有较高专一性,而且水解产率较高,所以可以选择各种不同专一性的酶进行专一性的断裂。
常用的酶有胰蛋白酶、糜蛋白酶、胃蛋白酶和嗜热菌蛋白酶。
胰蛋白酶专一断裂Lys,Arg的羧基侧肽键,如果对Lys,Arg,CysH进行化学修饰可改变胰蛋白酶的断裂性质。
(1)赖氨酸的修饰。将Lys用顺丁烯二酸酐或甲基顺丁烯二酸杆修饰,则胰蛋白酶仅使Arg肽键断裂。
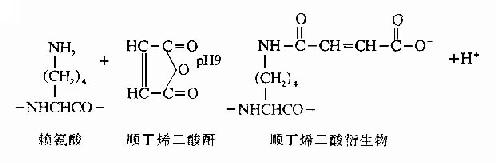
顺丁烯衍生物在中性pH下稳定,胰蛋白酶水解仅使Arg键断裂。在酸性条件下顺丁烯衍生物可脱去封闭,此时再行胰蛋白酶水解,则得赖氨酸为末端的多肽。下述为蛋白质中的赖氨酸,经顺丁烯酰化作用后,被胰蛋白酶水解的例子。
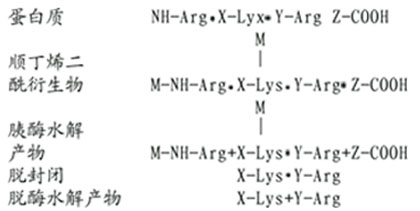
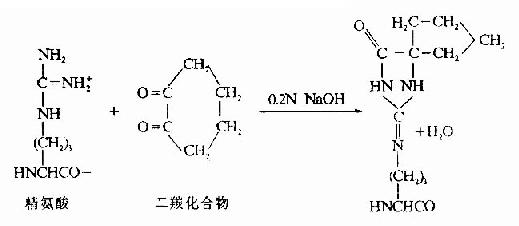
(2)精氨酸的修饰。精氨酸与1,2-和1,3-二羰化合物作用,缩合产物是一杂环化合物,十分稳定。然后胰酶水解仅断裂赖氨酸残基末端的肽键
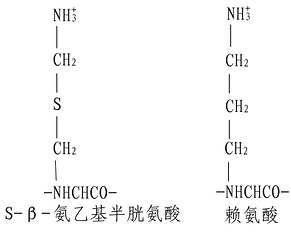
(3)半胱氨酸的修饰。若肽链内Lys、Arg均较少,则为了增加胰酶的裂解点,可以将半胱氨酸进行氨乙基化,其产物Sβ氨乙基半胱氨酸有类似Lys的结构,胰蛋白酶在水解时,不能识别这微细的变化,从而在半胱氨酸处断裂。
蛋白水解酶的专一性
| 酶 | 米源 | 主要作用点 | 其它作用点 |
| 胰蛋白酶 | 胰 | Arg,Lys | |
| 糜蛋白酶 | 胰 | Tyr,Phe,Trp | Leu,Met,His,Asu,Gln |
| 弹性蛋白酶 | 胰 | Leu,Ile,Ala | 其它等 |
| 胃蛋白酶 | 胃粘膜 | Tyr,Phe,Trp,Met,Len | Ala,Glu,Asp,其它等 |
| 木瓜蛋白酶 | Papayplant木瓜植物 | Arg,Lys,Gly | 其它等 |
| 嗜热菌蛋白酶 | 嗜热解蛋白芽孢杆菌 | Leu,Ile,Phe | Val,Tyr |
| 枯草杆菌蛋白酶 | 枯草杆菌 | 芳香族及脂肪族残基 |
肽链的裂解和重组大致有三种情况:一种是非特异性裂解,如酸水解。由于裂解的片段较小,造成分离的困难。因此这种非特异性裂解对大分子肽链是不适用的。第二种是特异性裂解,采用两种以上的专一裂解,然后进行组合,这种方法一般也适用于分子量小于5万的蛋白质。第三种是逐步的专一裂解,首先将某种氨基酸进行化学修饰,使水解酶专一断裂某一种氨基酸,分成若干片段,然后解除化学封闭,再用此酶裂解,使曾被封闭过的氨基酸断裂。目前倾向于采用这种裂解方式。
3.肽段的分离
大部分肽段的分离主要通过凝胶过滤法,由于大分子肽溶解度小。往往采用甲酸、醋酸、丙酸等有机溶剂使之溶解。单用凝胶过滤法分离之肽一般纯度不高,常需辅以离子交换层析法,大片段肽可用离子交换葡聚糖作载体,小肽则多用Dowex-50等树脂。
小肽分离还常采用高压电泳与层析相结合的指纹图谱法,得到纯净肽。
4.肽的顺序分析
在蛋白质一级结构的测定中,肽的顺序分析是比较重要的一步。肽的顺序分析也有化学法和酶解法两种。
1)化学法Edman降解法
这是目前用于顺序分析的最主要的方法。它的原理是从N端开始,逐步降解。将肽先与异硫氰酸苯酯(PTH试剂)在pH8-9条件下作用,肽的NH2末端接到异硫氰酸苯酯的C原子上生成苯异硫甲氨酰肽,简称PTC肽,在强酸作用下,可使靠近PTC基的氨基酸环化,肽键断裂形成苯氨基噻唑啉酮衍生物和一个失去末端氨基酸的肽链。此肽不被破坏,因而又可出现一个新的N-末端。重复以上的步骤,继续与PTH试剂作用,继续分析,苯氨基噻唑啉酮衍生物很容易由有机溶剂抽提出来进行鉴定。但此衍生物很不稳定,在水中可转化为稳定的乙内酰苯硫脲氨基酸(PTH-氨基酸)。这些步骤通常称为Edman氏逐步降解法。所以可用来测定氨基酸的排列顺序。Edman降解法的优点是样品用量少,灵敏度高。

PTH-氨基酸的鉴定可以用各种层析方法,如纸层析、薄层层析、气相层析和质谱法等,现在多用高压液相层析法。虽然此方法具有很多优点,但是由于操作繁琐,工作量大,所以目前有人根据Edman降解的原理作一系列改进。
下面简单介绍几种方法
A.1967年Edman及Begg介绍了一种Edman降解的液相自动分析装置,使顺序分析开始走向自动化。将样品先在反应杯内旋转成薄膜,使之固定。然后与PITC试剂反应。再用有机溶剂多次抽提除去过剩试剂,因而样品易丢失,且仪器昂贵,使用受到限制。
B.1970年Laursen改进为固相氨基酸顺序仪。此法样品用量少,检出灵敏,可分析20?0肽,其原理是将肽共价结合到惰性支持物上,固定后装柱再行Edman降解。
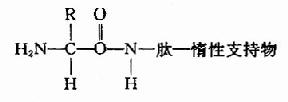
固相顺序仪的惰性支持物有:

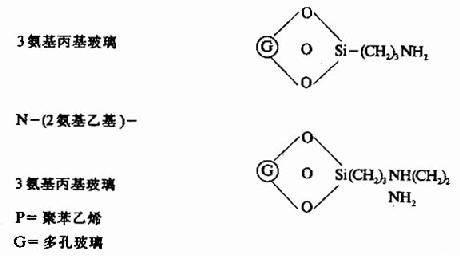
此法成功的关键是肽段的固定,目前采用C端α羧基固定法,重复法高,其中以高丝氨酸内酯法及双异硫氰酯法(DITC)最好,固定率可达90-95%。
C.另外也有从化学反应的角度考虑,试图改进Edman方法。1976年有人将异硫氰酸苯酯的苯基改变为甲氨偶氮苯,试剂为甲氨偶氨苯异硫氰酸盐(简称DABITC)。这是一种有色试剂,产物DABIH氨基酸呈桔黄色,因此鉴定时无需染色,用肉眼即可分辨。此方法灵敏度很高,一次分析小肽段只要几个nanomole样品即可,是目前一种很可取的方法。此外也有人将异硫氰酸酯进行35S标记,使分析样品更向微量化方向发展。
2)酶解法肽谱重迭法
分析肽段也可采用酶解法,利用专一性不同的两种酶将一个肽分别断裂成更小的寡肽,比较两种方法所得之肽段的重复性,进行氨基酸顺序的装配。例如,有一个肽段,通过氨基酸组成分析已知其为十肽,假如先以糜蛋白酶水解,则得到一套寡肽,再以胰蛋白酶水解此十肽,得到另一套寡肽。分析结果如下:
Ala·Phe+Gly·Lys·Asn·Tyr+Arg·Trp+His·Val
糜蛋白酶水解
+肽(Ala·Phe·Gly·Lys·Asn·Tyr·Arg·Trp·His·Val)
胰蛋白酶水解
Ala·Phe·Gly·Lys+Asa·Tyr·Arg+Trp·His·Val
将此两套寡肽可以做分析比较,因为十肽的N末端及C末端已事先测定分别为Ala及Val,因此第一段寡肽必然是Ala,Phe。如此类推如下
| 寡肽号 | 氨基酸组成部分顺序 |
| A-1 | Ala·Pha |
| B-1 | Ala·Phe·Gly·Lys |
| A-2 | Gly·Lys·Asn·Tyr |
| B-2 | Asn·Tyr·Arg |
| A-3 | Arg·Trp |
| B-3 | Trp·His·Val |
| A-4 | His·Val |
| +肽顺序 | Ala·Phe·Gly·Lys·Asn·Tyr·Arg·Trp·His·Val |
水解酶也可运用二肽酶,两组可用同一种酶水解如第一套肽是A桞,C桪,E桭,G桯……第二套肽水解则先将该肽段N端切去一个末位氨基酸,然后再开始二肽酶断裂,结果是A,B桟,D桬,F桮……这样分析比较也可排列出肽段顺序。
5.二硫键定位
蛋白质分子不经任何处理,直接用酶水解,检出其中二硫键的肽段,然后将二硫键拆开,分别测定两个肽的顺序,将此两肽结构与测出的一级结构比较,就能找出相应的二硫键的位置。
含二硫键肽的检出方法。
1)凝胶过滤或离子交换层析:用以分离各肽段,然后用特殊的二硫键显色反应找出含二硫键的肽。
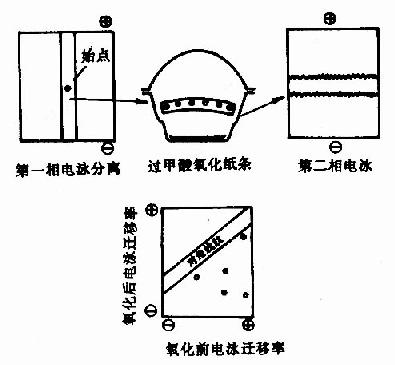
图1-21 对角线电泳技术图解
2)对角线电泳或层析:1966年Brown及Hartlay提出用对角线电泳进行含-S-S-肽的定位,此方法是将水解后的肽混合物进行第一相电泳,样品点在中间,电泳毕,将样品纸条剪下,置于装有过甲酸的器皿中,用过甲酸蒸气处理2小时,使-S-S-断裂,此时含-S-S-肽段的静电荷发生了改变。然后将纸条缝于另一张纸上,进行第二相电泳电泳,电泳条件与第一相相同,只是与第一次方向成直角。在第二相电泳中,那些不含-S-S-的髣民泳情况与第一相相同,因此电泳后各肽斑均坐落在纸的对角线上,而那些含-S-S-的肽由于被氧化,电荷发生变化,第二相电泳速度就与第一相不同,电泳结果这些肽斑就偏离对角线,肽斑可用茚三酮显示。对角线法由于其速度快,操作简便以及能用于小分子样品,是直接分离-S-S-肽的好方法。
含-S-S-肽被分离后,即可进行肽段顺序分析,并与已测定的该蛋白质的一级结构进行比较,即可找出相应的-S-S-位置,至此蛋白质的一级结构基本阐明。
今后蛋白质一级结构的测定正朝自动化、快速化及微量化发展,关键问题仍然是进一步寻找蛋白裂解和肽分离的方法。
蛋白质一级结构的测定不断有新方法和新思路出现,如X衍射法测定一级结构;分离相应蛋白质的mRNA,由mRNA的一级结构排出蛋白质的一级结构等。这些大胆的设想必将有助于蛋白质的一级结构测定,使人们掌握更多的工具和方法去探索生命的奥秘。
(毛积芳、刘新平)