第四节 呼吸运动的调节
呼吸运动是一种节律性的活动,其深度和频率随体内、外环境条件的改变而改变例如劳动或运动时,代谢增强,呼吸加深加快,肺通气量增大,摄取更多的O2,排出更多的CO2,以与代谢水平相适应。呼吸为什么能有节律地进行?呼吸的浓度和频率又如何能随内、外环境条件而改变?这些总是是本节的中心。
一、呼吸中枢与呼吸节律的形成
呼吸中枢是指中枢神经系统内产生和调节呼吸运动的神经细胞群。多年来,对于这些细胞群在中枢神经系统内的分布和呼吸节律产生和调节中的作用,曾用多种技术方法进行研究。如早期的较为粗糙的切除、横断、破坏、电刺激等方法,和后来发展起来的较为精细的微小电毁损、微小电刺激、可逆性冷冻或化学阻滞、选择性化学刺激或毁损、细胞外和细胞内微电极记录、逆行刺激(电刺激轴突,激起冲动逆行传导至胞体,在胞体记录)、神经元间电活动的相关分析以及组织化学等方法。有管些方法对动物呼吸中枢做了大量的实验性研究,获得了许多宝贵的资料,形成了一些假说或看法。
(一)呼吸中枢
呼吸中枢分布在大脑皮层、间脑、脑桥、延髓和脊髓等部位。脑的各级部位在呼吸节律产生和调节中所起作用不同。正常呼吸运动是在各级呼吸中枢的相互配合下进行的。
1.脊髓 脊髓中支配呼吸肌的运动神经元位于第3-5颈段(支配膈肌)和胸段(支配肌间肌和腹肌等)前角。很早就知道在延髓和脊髓间横断脊髓,呼吸就停止。所以,可以认为节律性呼吸运动不是在脊髓产生的。脊髓只是联系上(高)位脑和呼吸肌的中继站和整合某些呼吸反射的初级中枢。
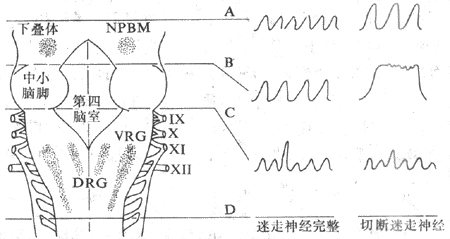
2.下(低)位脑干 下(低)位脑干指脑桥和延髓。横切脑干的实验表明,呼吸节律产生于下位脑干,呼吸运动的变化因脑干横断的平面高低而异(图5-17)。
图5-17脑干呼吸有关核团(左)和在不同平面横切脑干后呼吸的变化(右)示意图
DRG:背侧呼吸组 VRH:腹侧呼吸组 NPBM:臂旁内侧核
A、B、C、D为不同平面横切
在动物中脑和脑桥之间进行横切(图5-17,A平面),呼吸无明显变化。在延髓和脊髓之间横切(D平面),呼吸停止。上述结果表明呼吸节律产生于下位禽干,上位脑对节律性呼吸不是必需的。如果在脑桥上、中部之间横切(B平面),呼吸将变慢变深,如再切断双侧迷走神经,吸气便大大延长,仅偶尔为短暂的呼气所中断,这种形式的呼吸称为长吸呼吸。这一结果是提示脑桥上部有抑制吸气的中枢结构,称为呼吸整中枢;来自肺部的迷走传入冲动也有抑制吸气的作用,当延髓失去来自这两方面对吸气活动的抑制作用后,吸气活动不能及时中断,便出现长吸呼吸。再在脑桥和延髓之间横切(C平面),不论迷走神经是否完整,长吸式呼吸都消失,而呈喘息样呼吸,呼吸不规则,或平静呼吸,或两者交替出现。因而认为脑桥中下中有活化吸气的长吸中枢;单独的延髓即可产生节律呼吸。孤立延髓的实验进一步证明延髓可独立地产生节律呼吸。于是在20-50年代期间形成了三级呼吸中枢理论;脑桥上部有呼吸调整中枢,中下部有长吸中枢,延髓有呼吸节律基本中枢。后来的研究肯定了早期关于延髓有呼吸节律基本中枢和脑桥上部有呼吸调整中枢的结论,但未能证实脑桥中下部存在着结构上特定的长吸中枢。
近年来,用微电极等新技术研究发现,在中枢神经系统内有的神经元呈节律性放电,并和呼吸周期相关,这些神经元被称为呼吸相关神经元或呼吸神经元。这些呼吸神经元有不同类型。就其自发放电的时间而言,在吸气相放电的为吸气神经元,在呼气相放电的为呼气神经元,在吸气相放电并延续至呼气相的为吸气-呼气神经元,在呼气相放电并延续到吸气相者,为呼气-吸气神经元,后两类神经元均系跨时相神经元。
在延髓,呼吸神经元内主要集中在背侧(孤束核的腹外侧部)和腹侧(疑核、后疑核和面神经后核附近的包氏复合体)两组神经核团内,分别称为背侧呼吸组(dorsal respiratory group,DRG)和腹侧呼吸组(ventral respiratory group,VRG)(图5-17)。背侧呼吸组的神经元轴突主要交叉到对侧,下行至脊髓颈段,支配膈运动神经元。疑核呼吸神经元的轴突由同侧舌咽神经和迷走神经传出,支配咽喉部呼吸辅助肌。后疑核的呼吸神经元绝大部分交叉到对侧下行,支配脊髓肌间内、外肌和腹肌的运动神经元,商品化纤维也发出侧支支配膈肌的运动神经元。包氏复合体主要含呼气神经元,它们的轴突主要与背侧呼吸组的吸气神经元形成抑制性联系,此外也有轴突支配脊髓的膈运动神经元。
由于延髓呼吸神经元主要集中在背侧呼吸组和腹侧呼吸组,所以曾推测背侧呼吸组和腹侧呼吸组是产生基本呼吸节律的部位。可是,后来的某些实验结果不支持这一看法。有人用化学的或电解的毁损这些区域后,呼吸节律没有明显变化,这些结果提示背侧呼吸组和腹侧呼吸组可能不是呼吸节律唯一发源地,呼吸节律可能源于多个部位,产生呼吸节律的神经结构相当广泛,所以不容易因局灶损害而丧失呼吸节律。
在脑桥上 部,呼吸神经元相对集中于臂旁内侧核和相邻的Kolliker-Fuse(KF)核,合称PBKF核群。PBKF和延髓的呼吸神经核团之间有双向联系,形成调控呼吸的神经元回路。在麻醉猫,切断双侧迷走神经,损毁PBKF可出现长吸,提示早先研究即已发现的呼吸调整中枢乃位于脑桥的BPKF,其作用为限制吸气,促使吸气向呼气转换。
3.上位脑 呼吸还受脑桥以上部位的影响,如大脑皮层、边缘系统、下丘脑等。
大脑皮层可以随意控制呼吸,发动说、唱等动作,在一定限度内可以随意屏气或加强加快呼吸。大脑皮层对呼吸的调节系统是随意呼吸调节系统,下位脑干的呼吸调节系统是自主节律呼吸调节系统。这两个系统的下行通路是分开的。临床上有时可以观察到自主呼吸和随意呼吸分离的现象。例如在脊髓前外侧索下行的处主呼吸通路受损后,自主节律呼吸甚至停止,但病人仍可进行随意呼吸。患者靠随意呼吸或人工呼吸来维持肺通气,如未进行人工呼吸,一旦病人入睡,可能发生呼吸停止。
(二)呼吸节律形成的假说
呼吸节律是怎样产生的,尚未完全阐明,已提出多种假说,当前最为流行的是局部神经元回路反馈控制假说。
中枢神经系统里有许多神经元没有长突起向远处投射,只有短突起在某一部位内形成局部神经元回路联系。回路内可经正反馈联系募集更多神经元兴奋,以延长兴奋时间或加强兴奋活动;也可以负反馈联系,以限制其活动时间或终止其活动。平静呼吸时,由于吸气是主动的,所以许多学者更多地是去研究吸气中如何发生的,又如何转变为呼气的。有人提出中枢吸气活动发生器和吸气切断机制(inspiratory off-switch mechanism)的看法,认为在延髓有一个中枢吸气活动发生器,引发吸气神经元呈斜坡样渐增性放电,产生吸气;还有一个吸气切断机制,使吸气切断而发生呼气。在中枢吸气活动发生器作用下,吸气神经元兴奋,其兴奋传至①脊髓吸气肌运动神经元,引起吸气,肺扩张;②脑桥臂旁内侧核,加强其活动;③吸气切断机制,使之兴奋。吸气切断机制接受来自吸气神经元,脑桥背旁内侧核,和肺牵张感觉器的冲动。随着吸气相的进行,来自这三方面的冲动均逐渐增强,在吸气切断机制总合达到阈值时,吸气切断机制兴奋,发出冲动到中枢吸气活动发生器或吸气神经元,以负反馈形式终止其活动,吸气停止,转为呼气(图5-18)。切断迷走神经或毁损脑桥臂旁内侧核或两者,吸气切断机制达到阈值所需时间延长,吸气因面延长,呼吸变慢。因此,凡可影响中枢吸气活动发生器、吸气切断机制阈值或达到阈值所需时间的因素,都可影响呼吸过程和节律。
关于呼气如何转入吸气,呼吸加强时呼气又如何成为主动的,目前了解料少。
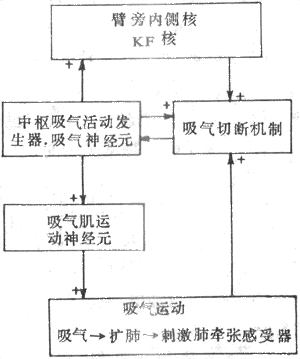
图5-18 呼吸节律形成机制简化模式图
+:表示兴奋 -:表示抑制
二、呼吸的反射性调节
呼吸节律虽然产生于脑,但其活动可受来自呼吸器官本身以及骨骼肌、其它器官系统感觉器‘传入冲动的反射性调节,下述其中的一些重要反射
(一)肺牵张反射
1868年Breuer和Hering发现,在麻醉动物肺充气或肺扩张,则抑制吸气;肺放气或肺缩小,则引起吸气。切断迷走神经,上述反应消失,所以是反射性反应。由肺扩张或肺缩小引起的吸气抑制或兴奋的反射为黑-伯反射(Hering-Breuer reflex)或肺牵张反射。它有两种成分:肺扩张反射和肺缩小反射。
1.肺扩张反射 是肺充气或扩张时抑制吸气的反射。感觉器位于从气管到细支气管的平滑肌中,是牵张感受器,阈值低,适应慢。当肺扩张牵拉呼吸道,使之也扩张时,感觉器兴奋,冲动经迷走神经走神经粗纤维传入延髓。在延髓内通过一定的神经联系使吸气切断机制兴奋,切断吸气,转入呼气。这样便加速了吸气和呼气的交替,使呼吸频率增加。所以切断迷走神经后,吸气延长、加深,呼吸变得深而慢。
有人比较了8种动物的肺扩张反射,发现有种属差异,兔的最强,人的最弱。在人体,当潮气量增加至800ml以上时,才能引起肺扩张反射,可能是由于人体肺扩张反射的中枢阈值较高所致。所以,平静呼吸时,肺扩张反射不参与人的呼吸调节。但在初生婴儿,存在这一反射,大约在出生4-5天后,反射就显著减弱。病理情况下,肺顺应性降低,肺扩张时使气道扩张较大,刺激较强,可以引起该反射,使呼吸变浅变快。
2.肺缩小反射 是肺缩小时引起吸气的反射。感受器同样位于气道平滑肌内,但其性质尚不十分清楚。肺缩小反向在较强的缩肺时才出现,它在平静呼吸调节中意义不大,但对阻止呼气过深和肺不张等可能起一定作用。
(二)呼吸肌本体感受性反射
肌梭和腱器官是骨骼肌的本体感受器,它们所引起的反射为本体感受性反射。如肌梭受到牵张刺激时可以反射性地引起受刺激肌梭所在肌的收缩,为牵张反射,属本体感受性反射(参见第十章第四节)。呼吸肌也有牵张反射的主要依据是:在麻醉猫,切断双侧迷走神经,颈7横断脊髓,牵拉膈肌,膈肌肌电活动啬;切断动物的胸脊神经背根,呼吸运动减弱;人类为治病需要曾做类似手术,术后相应呼吸肌的活动发生可恢复的或可部分恢复的减弱。说明呼吸肌本体感受性反射参与正常呼吸运动的调节,在呼吸肌负荷改变时将发挥更大的作用。但是,这些依据不是无懈可击的。因为背根切断术不仅切断了本本感受器的传入纤维,也切断了所有经背根传入的其它感受器的传入纤维。近来的研究表明来自呼吸肌其它感受器的传入冲动也可反射性地影响呼吸。因此,对呼吸肌本体感受性反射应做更深更深入细致的研究,如研究分别兴奋不同感受器或传入纤维时对呼吸的效应。
(三)防御性呼吸反射
在整个呼吸道都存在着感受器,它们是分布在粘膜上皮的迷走传入神经末梢,受到机械或化学刺激时,引起防御性呼吸反射,以清除激惹物,避免其进入肺泡。
1.咳嗽反射 是常见的重要防御反射。它的感受器位于喉、气管和支气管的粘膜。大支气管以上部位的感受器对机械刺激敏感,二级支气管以下部位的对化学刺激敏感。传入冲动经迷走神经传入延髓,触发一系列协调的反射反应,引起咳嗽反射。
咳嗽时,先是短促或深吸气,接着声门紧闭,呼气肌强烈收缩,肺内压和胸膜腔内压急速上升,然后声门突然打开,由于气压差极大,气体更以极高的速度从肺内冲出,将呼吸道内异物或分泌物排出。剧烈咳嗽时,因胸膜腔内压显著升高,可阻碍静脉因流,使静脉压和脑脊液压升高。
2.喷嚏反射 是和咳嗽类似的反射,不同的是:刺激作用于鼻粘膜感受器,传入神经是三叉神经,反射效应是腭垂下降,舌压向软腭,而不是声门关闭,呼出气主要从鼻腔喷出,以清除鼻腔中的刺激物。
(四)肺毛细血管旁(J-)感受器引起的呼吸反射
J-感受器位于肺泡毛细血管旁,在肺毛细血管充血、肺泡壁间质积液时受到刺激,冲动经迷走神经无髓C纤维传入延髓,引起反射性呼吸暂停,继以浅快呼吸,血压降低,心率减慢。J-感受器在呼吸调节中的作用尚不清楚,可能与运动时呼吸加快作肺充血、肺水肿时的急促呼吸有关。
(五)某些穴位刺激的呼吸效应
针刺人中窕可以急救全麻手术过程中出现的呼吸停止。针刺动物人中可以使膈肌呼吸运动增强,电刺激家兔人中对膈神经和管髓呼吸神经元电活动有特异性影响。有人观察到在麻醉意外事件发生呼吸暂停时,刺激素可以兴奋呼吸。穴位的呼吸效应及其机制值得探讨。
(六)血压对呼吸的影响
血压大幅度变化时可以反射性地影响呼吸,血压升高,呼吸减弱减慢;血压降低,呼吸加强加快。
三、化学因素对呼吸的调节
化学因素对呼吸的调节也是一种呼吸的反射性调节,化学因素是指动脉血或脑脊液中的O2、CO2和H+。机体通过呼吸调节血液中的O2、CO2和H+的水平,动脉血中O2、CO2和H+水平的变化又通过化学感受器调节着呼吸,如此形成的控制环维持着内环境这些因素的相对稳定。
(一)化学感受器
化学感觉器是拂晓春适宜刺激化学物质的感受器。参与呼吸调节的化学感受器因其所在部位的不同,分为外周化学感受器和中枢化学感受器。
1.外周化学感受器 颈动脉体和主动脉体是调节呼吸和循环的重要外周化学感受器。在动脉血PO2降低、PCO2或H+浓度([H+])升主时受到刺激,冲动经窦神经和迷走神经传入延髓,反射性地引起呼吸加深加快和血液循环的变化。虽然颈、主动脉体两者都参与呼吸和循环的调节,但是颈动脉体主要调节呼吸,而主动脉体在循环调节方面较为重要。由于颈动脉体的有利的解剖位置,所以,对外周化学感受器的研究主要集中在颈动脉体。
颈动脉体含Ⅰ型细胞(球细胞)和Ⅱ型细胞(鞘细胞),它们周围包绕以毛细血管窦。血液供应十分丰富。Ⅰ型细胞呈球形,有大量囊泡,内含递质,如乙酰胆碱、儿茶酚胺、某些神经活性肽等。Ⅱ型细胞数量较少,没有囊泡。Ⅱ型细胞包绕着Ⅰ型细胞、神经纤维和神经末梢,功能上类似神经胶质细胞,与颈动脉体其它成分之间没有特化的接触。窦神经的传入纤维末梢分支穿插于Ⅰ、Ⅱ型细胞之间,与Ⅰ型细胞形成特化接触,包括单向突触、交互突触、缝隙边接等(图5-19),传入神经末梢可以是突触前和(或)突触后成分。交互突触构成Ⅰ型细胞与传入神经之间的一种反馈环路,借释放递质调节化学感受器的敏感性。此外,颈动脉体还有传出神经支配,借调节血流和化学感受器以改变化学感受器的活动。
用游离的颈动脉体,记录其传入神经单纤维的动作,观察改变灌流液成分时动作频率的变化,可以了解颈动脉体所感受的刺激的性质以及刺激与反应之间的关系。结果发现当灌流液PO2下降,PCO2或[H+]升高时,传入冲动增加。如果保持灌流血液的PO2正常的13.3kPa(100mlHg),仅减少血流量,传入冲动也增加。困为血流量下降时,颈动脉体从单位血液中摄取的O2量相对增加,细胞外液 PO2因供O2少于耗 O2而下降。但在贫血或CO中毒时,血 O2含量虽然下降,但PO2正常,只需血流量充分,化学感受器传入冲动并不增加,所以化学感受器所感受的刺激是PO2,而不是动脉血O2含量,而且是感受器所处环境的PO2。从实验中还可看出上述三种刺激对化学感受器有相互增强的作用。两种刺激同进作用时比单一刺激的效应强。这种协同作用有重要意义,因为机体发生循环或呼吸衰竭时,总是PCO2升高和PO2降低同进存在,它们的协同作用加强了对化学感受器的刺激,从而促进了代偿性呼吸增强的反应。
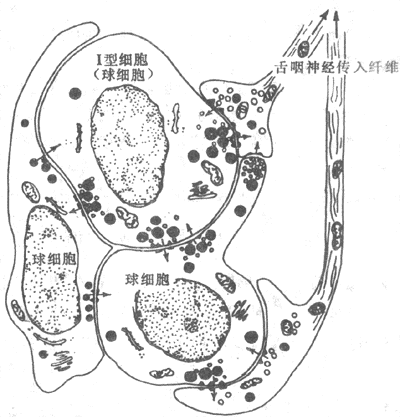
图5-19 颈动脉体组织结构示意图
图中未显示Ⅱ细胞
目前认为,Ⅰ型细胞起着化学感受器的作用。当它们受到刺激时,细胞浆内[Ca2+]升高。触发递质释放,引起传入神经纤维兴奋。PO2降低与 PCO2或[H+]升高引起细胞内[Ca2+]升高机制不同。PO2降低可抑制细胞 K+通道的开放,K+外流减少,细胞膜去极化,从而促使电压依从性Ca2+通道开放,Ca2+进入细胞。而PCO2或[H+]升高时,进入细胞内的H+增多,激活了细胞的Na+-H+交换机制,Na+进入细胞,使细胞内[Na+]长高,继而使细胞的Na+-Ca2+交换机制活动啬,Na+出细胞,Ca2+进细胞内,引起细胞浆内[Ca2+]升高。还有资料表明,少部分胞浆内Ca2+可能来自细胞内的Ca2+贮器。
2.中枢化学感受器 摘除动物外周化学感受器或切断其传入神经后,吸入CO2仍能加强通气。改变脑脊液CO2和H+浓度也能刺激呼吸。过去认为这是CO2直接刺激呼吸中枢所致年代以来,用改变脑表面灌流液成分和pH、局部冷阻断、电凝固损伤、电刺激、记录神经元电活动、离体脑组织块的电生理研究等方法在多种动物做了大量实验,结果表明在延髓有一个不同于呼吸中枢,但可影响呼吸的化学感受器,称为中枢化学感受器,以另于外周化学感受器。
中枢化学感受器 位于延髓腹外侧浅表部位,左右对称,可以分为头、中、尾三个区(图5-20A)。头端和尾端区都有化学感受性,中间区不具有化学感受性,不过,局部阻滞或损伤中间区后,可以使动物通气量降低,并使头端、尾端区 受刺激时的通气反应消失,提示中间区可能是端区和尾 端区传入冲动向脑干呼吸中枢投射的中继站。应用胆碱能激动剂和拮抗剂的研究结果表明,在中枢化学感受器传递环节中可能有胆碱能机制参与。
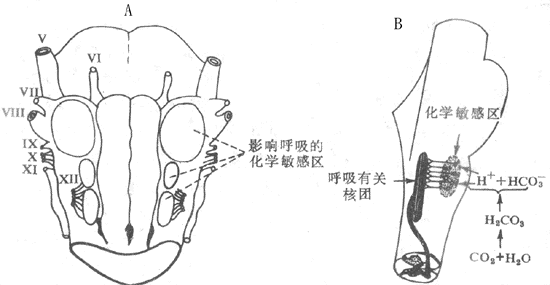
图5-20 中枢化学感受器
A示延髓腹外侧的三个化学敏感区 B示血液或
脑脊液PCO2升高时,刺激呼吸的中枢机制
中枢化学感受器的生理刺激是脑脊液和局部细胞外液的H+。因为如果保持人工脑脊液的pH不变,用含高浓度CO2的人工脑脊液灌流脑室时所引起的通气增强反应消失,可见有效刺激不是CO2本身,而是CO2所引起的[H+]的增加。在体内,血液中的CO2能迅速通过血脑屏障,使化学感受器周围液体中的[H+]升高,从而刺激中枢化学感受器,再引起呼吸中枢的兴奋(图5-20B)。可是,脑脊液中碳酸酶含量很少,CO2与水的水合反应很慢,所以对CO2的反应有一定的时间延迟。血液中的H+不易以通过血液屏障,故血液pH的变化对中枢化学感受器的直接作用不大,也较缓慢。
中枢化学感受器与外周化学感受器不同,它不感受缺O2的刺激,但对CO2的敏感性比外周的主,反应潜伏期较长。中枢化学感受器的作用可能是调节脑脊液的[H+],使中枢神经系统有一稳定的pH环境,而外周化学感受器的作用主要是在机体低O2时,维持对呼吸的驱动。
(二)CO2、H+和O2对呼吸的影响
1.CO2的影响 很早已经知道,在麻醉动物或人,动脉血液PCO2降得很低时可发生呼吸暂停。因此,一定水平的PCO2对维持呼吸和呼吸中枢的兴奋性是必要的,CO2是调节呼吸的最重要的生理性体液因子。
吸入含CO2的混合气,将使肺泡气PCO2长高,动脉血PCO2也随之升高,呼吸加深加快,肺通气量增加(图5-21)。通过肺通气量的增大可能增加CO2的清除,肺泡气和动脉血PCO2还可维持于接近正常水平。但是,当吸入气CO2陡升,CO2堆积,压抑中枢神经系统的活动,包括呼吸中枢,发生呼吸困难、头痛、头昏,甚至昏迷,出现CO2麻醉。对CO2的反应,有个体差异,还受许多因素影响,如疾病或药物。总之CO2在呼吸调节中是经常起作用的最重要的化学刺激,在一定范围内动脉血PCO2的升高,可以加强对呼吸的刺激作用,但超过一定限度则有压抑和麻醉效应。
CO2刺激呼吸是通过两条途径实现的,一是通过刺激中枢化学感受器再兴奋呼吸中枢:二是刺激外周化学感受器,冲动窦神经和迷走神经传入延髓呼吸有关疑团,反射性地使呼吸加深、加快,增加肺通气。但两条途径中前者是主要的。因为去掉外周化学感受器的作用之后,CO2的通气反应仅下降约20%,可见中枢化学感受器在CO2通气反应中起主要作用;动脉血PCO2只需升高 0.266kPa(2mmHg)就可刺激中枢化学感受器,出现通气加强反应,如刺激外周化学感受器,则需升高1.33kPa(10mmHg)。不过,在下述情况下,外周化学感受器的作用可能是重要的:因为中枢化学感受器的反应慢,所以当动脉血PCO2突然大增时,外周化学感受器在引起快速呼吸反应中可起重要作用;当中枢化学感受器到抑制,对CO2 的反应降低时,外周化学感受器就起重要作用。
2.H+的影响 动脉血[H+]增加,呼吸加深加快,肺通气增加;[H+]降低,呼吸受到抑制(图5-21)。H+对呼吸的调节也是通过外周化学感受器和中枢化学感受器实现的。中枢化学感受器对H+的敏感性较外周的高,约为外周的25倍。但是,H+通过血液屏障的速度慢,限制了它对中枢化学感受器的作用。脑脊液中的H+才是中枢化学感受器的最有效的刺激。
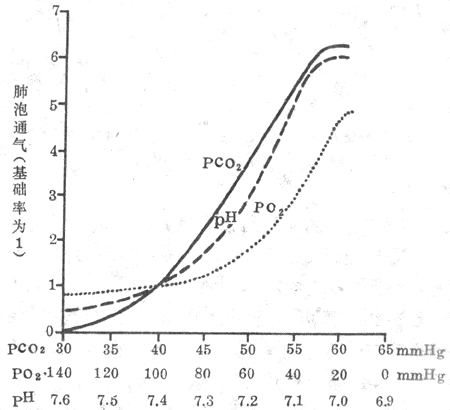
图5-21 动脉血液PCO2、PO2、pH改变对肺泡通气
的影响仅改变一种体液因素而保持另二因素于
正常水平时的情况(1mmHg=0.133kPa)
3.O2的影响 吸入气PO2降低时,肺泡气PO2都随之降低,呼吸加深、加快,肺通气增加(图5-21)。同CO2一样,对低O2的反应也有个体差异。一般在动脉PO2下降到10.64kPa(80mmHg)以下时,肺通气才出现可觉察到的增加,可见动脉血PO2对正常呼吸的调节作用不大,仅在特殊情况下低O2刺激才有重要意义。如严重肺气肿、肺心病患者,肺换气受到障碍,导致低O2和CO2潴留。长时间CO2潴留使中枢化学感受器对CO2的刺激作用发生适应,而外周化学感受器对低O2刺激适应很慢,这时低O2对外周化学感受器的刺激成为驱动呼吸的主要刺激。
低O2对呼吸的刺激作用完全是通过外周化学感受器实现的。切断动物外周化学感受器的传入神经或摘除人的颈动脉体,急性低O2的呼吸刺激反应完全消失。低O2对中枢的直接作用是压抑作用。但是低O2可以通过对外周化学感受器的刺激而兴奋呼吸中枢,这样在一定程度上可以对抗低O2对中枢的直接压抑作用。不过在严重低O2时,外周化学感受性反射已不足以克服低O2对中枢的压抑作用,终将导致呼吸障碍。在低O2时吸入纯O2,由于解除了外周化学感受器的低O2刺激,会引起呼吸暂停,临床上给O2治疗时应予以注意。
(三)PCO2、H+和PO2在影响呼吸中的相互作用
图5-21示保持其它两个因素不变而只改变其中一个因素时的单因素通气效应。可以看出PO2下降对呼吸的影响较慢、较弱,在一般动脉血PO2变化范围内作用不大,要在PO2低于10.64kPa(80mmHg)后,通气量才逐渐增大。PCO2和H+与低O2不同,只要略有升高,通气就明显增大,PCO2的作用尤为突出。

图5-22 动脉血液PCO2升高、PCO2降低、pH降低对肺泡通气率的影响
(1mmHg=0.133kpa)
但实际情况不可能是单因素的改变,而其它因素不变。往往是一种因素的改变会引起其余一、两种因素相继改变或存在几种因素的同时改变,三者间相互影响、相互作用,既可因相互总和而加大,也可因相互抵消而减弱。图5-22为一种因素改变,另两种因素不加控制时的情况。可以看出:PCO2升高时,[H+]也随之升高,两者的作用总和起来,使肺通气较单独PCO2升高时为大。[H+]增加时,因肺通气增大使CO2排出,PCO2下降,抵消了一部分H+的刺激作用;CO2含量的下降,也使[H+]有所降低。两者均使肺通气的增加较单独[H+]升高时为小。PO2下降时,也因肺通气量增加,呼出较多的CO2,使PCO2和[H+]下降,从而减弱了低O2的刺激作用。
四、周期性呼吸
周期性呼吸是异常呼吸型之一,表现为呼吸加强加快与减弱减慢交替出现。最常见的有陈-施呼吸和比奥(Biot)呼吸。
(一)陈-施呼吸(潮式呼吸)
陈-施呼吸(Cheyne-Stokes respiration)的特点是呼吸逐渐增强增快又逐渐减弱减慢与呼吸暂停交替出现,每个周期约45s到3min。
当前认为陈-施呼吸产生的基本机制是因为某种原因呼吸受到刺激,肺通气量增加,呼出过多的CO2,肺泡气PCO2下降,肺部血液PCO2也下降,片刻之后,这种低PCO2血液到达脑部,呼吸因缺少CO2的刺激而开始受到抑制,变慢变浅甚至停止。呼吸的抑制又使肺部血液PCO2升高,PCO2升高了的血液随后到达脑,又开始刺激呼吸,呼吸又复变快变深,再次使PCO2下降,呼吸再受抑制。上述过程周而复始,周期性进行,产陈-施呼吸(图5-23)。陈-施呼吸主要出现于二种情况下:①肺-脑循环时延长(如心力衰竭),此时脑PCO2将升高,增强了对呼吸的刺激,触了陈-施呼吸;②呼吸中枢反馈增益增加。反馈增益是指一定程度的PCO2或pH变化所引起的通气变化,通气变化大,则增益大。低O2或某种脑干损伤可出现增益增大,导致陈-施呼吸。
(二)Biot呼吸
其特点是一次或多次强呼吸后,继以长时间呼吸停止,之后又出现第二次这样的呼吸(图5-23)。周期持续时间变化较大,短的仅10s,长的可达1min。Biot呼吸见于脑损伤、脑脊液的压力升高、脑膜炎等疾病时,常是死亡前出现的危急症状。发生的原因尚不清楚,可能是疾病已侵及延髓,损害了呼吸中枢。
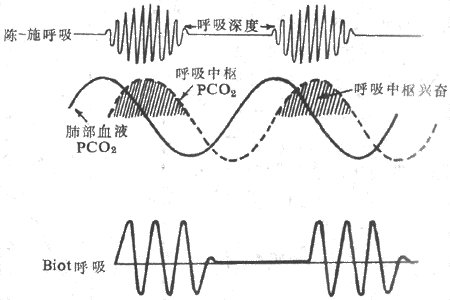
图5-23 异常呼吸型式示意图
五、运动时呼吸的变化及调节
运动时机体代谢增高,血液循环和呼吸系统都将发生一系列变化以适应增高了的机体代谢的需要。这时,呼吸加深加快,肺通气量增大,其增加的程度随运动量而异。潮气量可双安静时的500ml升到2000ml,呼吸频率可从 12-18次/min升至50次/min,每分通气量可升达100L以上,O2的摄入量和CO2排出量也都相应增加。
运动时肺通气量的增加有一个过程。运动之始,通气量骤升,继之以缓慢的升高,随后达一稳态水平。运动停止时,也是通气量先骤降,继以缓慢下降,然后恢复到运动前的水平(图5-24)。
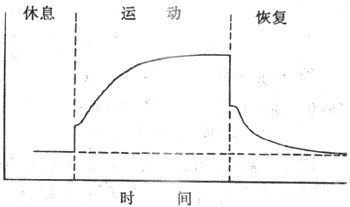
图5-24 运动时的肺通气量变化
运动时呼吸的变化是大家都很熟悉的生理现象,但其变化机制却至今仍未阐明,不过与其它生理活动一样,也是在神经和体液机制调节下发生的。
一般认为运动开始时通气骤升也条件反射有关,是在运动锻炼过程中形成的。因为只是给予运动暗示,并未开始运动,也可出现通气量增大的反应,而且与运动者过去的经验、精神状态、实验条件等有关。此外,运动时,运动肌肉、关节的本体感受器受到刺激,其传入冲动也可以反射性地刺激呼吸。因为,仅被动运动肢体,就可引起快速通气反应,阴断活动肢体的传入神经,反应消失;脊髓胸12水平截瘫病人,被动运动膝关节,不能产生快速通气反应。近年来神经组化研究表明,在延髓孤束周围有较稠密的肌传入末梢,这可能是运动肢体引起运动初期快速通气反应的解剖学基础。
运动时动脉血pH、PCO2PO2的波动的作用。中度运动时,虽然动脉血pH、PCO2PO2的均值保持相对的稳定,但它们却都能随呼吸厕于周期性波动,波动的幅度随运动强度而变化,运动强,波动幅度大;运动弱,波动幅度小。运动时,这种波动幅度的增大,可能在运动通气反应中起重要作用。动物实验中,设法在不影响血液气体平均分压的同时,缓冲上述周期性波动,动物通气量下降。记录猫颈动脉窦神经化学感受器传入冲动,发现其频率与呼吸周期同步消、涨。体液因素的这种作用是通过化学感受器实现的。
运动停止后,通气未立即恢复到安静水平。这是因为运动时,O2供小于O2耗,欠下了“O2债”。所以运动停止后,一段时间内,O2耗仍大于安静时的,以偿还O2债,待偿还后,通气才恢复。这时维持通气增强的刺激主要是H+。
参考资料
1.陈子彬延髓中枢化学感受器及其在呼吸调节中的作用,生理科学进展1982;13:300-305
2.张衡 关于哺乳动物呼吸中枢的现代观点生理科学进展 1986;17:210-215
3.郑煜 哺乳动物呼吸节律研究简介 生物学通报 1992;10:11-14
4.Numm JF,陈毓槐等译 应用呼吸生理学北京科学出版社,1987
5.Garby L,Meldon J,周殿松译 血液呼吸功能 北京 科学出版社 1987
6.Barnes PJ.Modulation of neurotransmission in airways.Physiol Rey 1992;72:699-729
7.Hay DWP,Henry PJ,Goldie RG.Endothelin and the respiratory system.TIPs 1993;14:29-32
8.gonzalez C,Almaraz L,Obeso A,Rigual R.Oxygen and acid chemoreception in the carotid body chemoreceptors .TINS1992;15:146-153
9.Cohen MI.Neurogenesis of respiratory rhythm in the mammal.Physiol Rev 1979;59:1105-1173
10.Slonim NB,Hamilton LH.Respiratory Physiology 5th ed,Cv Mosby Co,St Louis,1987